Gastrointestinal (GI) motility is the contractile activity responsible for mixing GI contents and for controlled propulsion of the contents distally.
Types of Muscle in the Gastrointestinal System
Striated muscle is found in the mouth, pharynx, upper esophagus, and external anal sphincter.
The remaining muscle tissue in the GI system is either circular or longitudinal smooth muscle, except in the stomach and gallbladder. The stomach has a layer of oblique muscle in addition to having circular and longitudinal muscle. The gallbladder is composed of smooth muscle, but the fibers are arranged in a reticulum (mesh).
Types of Contractions of Gastrointestinal Smooth Muscles
Phasic Contractions. Phasic contractions are cyclic contractions that permit mixing and propelling of GI contents. These types of contractions are important in the stomach and small intestine.
Tonic Contractions. Tonic contractions are continuous contractions that relax only under neural stimulation. The upper region of the stomach and the sphincters that control the flow of GI contents from one region of the GI tract to another demonstrate tonic contraction.
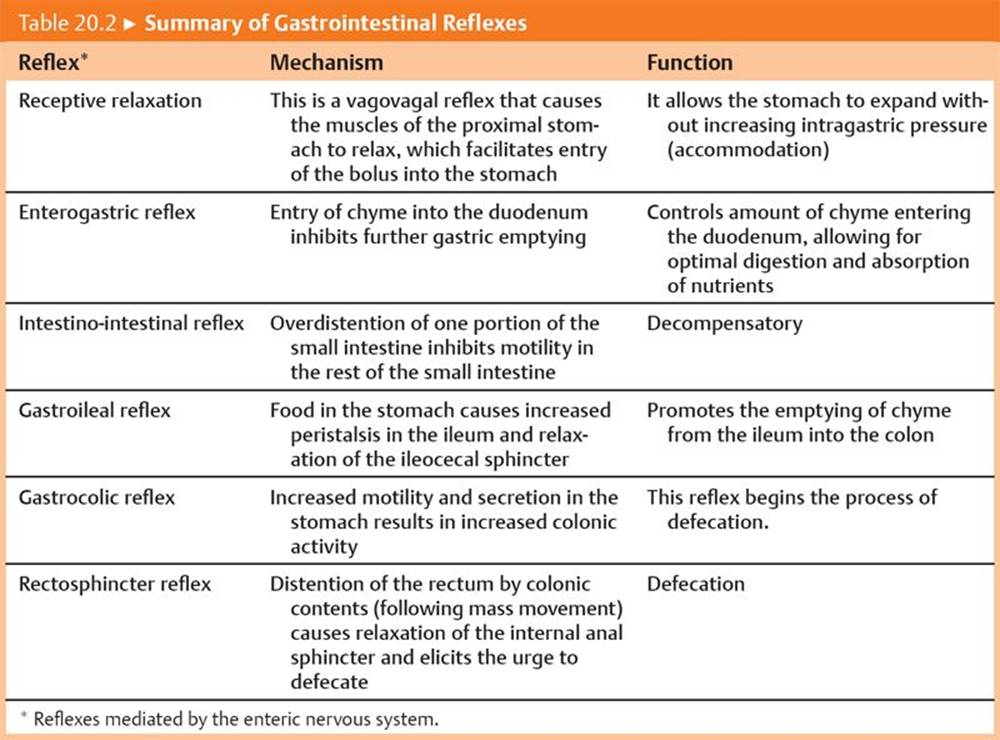
Slow Waves
Slow waves are oscillating waves of membrane depolarization that are not sufficient to completely depolarize the membrane and stimulate contractions, but that make it possible for contractions to be stimulated more easily by raising the membrane potential closer to threshold (making it less negative) (Fig. 20.1).
– Slow waves that act as pacemakers originate in the interstitial cells of Cajal.
– They are a feature of smooth muscle cells in the stomach, small intestine, and large intestine.
Frequency of Slow Waves
Slow waves determine the basic frequency of contractions throughout the GI system by facilitating membrane depolarization at a particular rate.
– Slow-wave frequency is modified by neural or hormonal activity. These influence the amplitude of the oscillation more than the frequency.
– The rate of slow waves is lowest in the stomach (3/min) and highest in the duodenum (12/min).
Fig. 20.1 ![]() Slow waves and spikes.
Slow waves and spikes.
The intestine contains pacemaker cells (interstitial cells of Cajal) that communicate with smooth muscle cells. The membrane potential of smooth muscle cells oscillates between 10 and 20 mV every 3 to 15 seconds, producing slow waves (1). These slow waves make the membrane potential less negative and, when the threshold potential is reached (~ 40 mV), a series of action potentials (spike bursts) are fired, resulting in muscle contraction (2). Muscle spasms occur if the trough of the wave also rises above the threshold potential (3). The smooth muscle cells are nonexcitable when they are depolarized (~ –20 mV) or hyperpolarized (~ –70 mV).
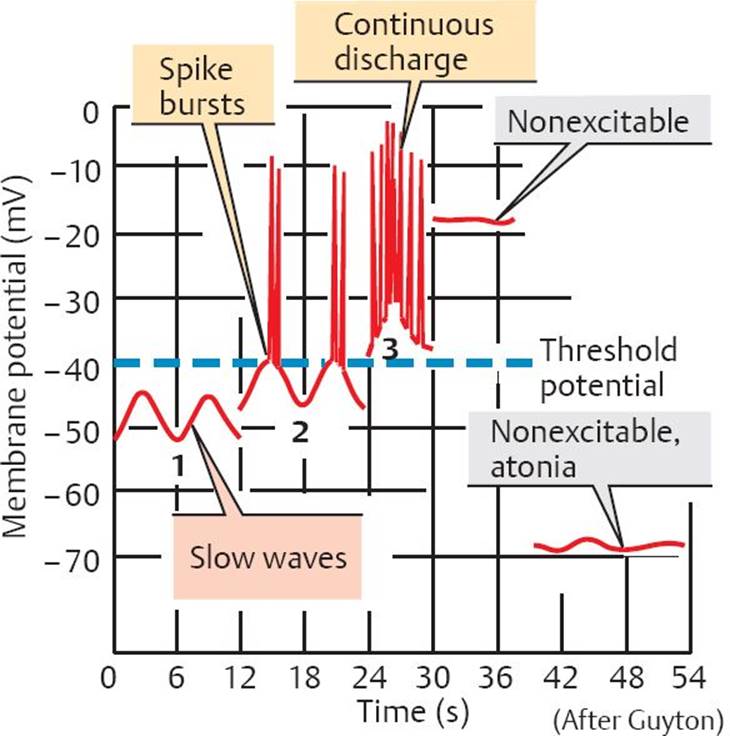
Fig. 20.2 ![]() Peristaltic reflex.
Peristaltic reflex.
Stretching of the intestinal wall during the passage of a bolus triggers a reflex that simultaneously contracts the circular muscles behind the bolus and relaxes the circular muscles in front of it. At the same time, longitudinal muscles behind the bolus are relaxed, and those in front of it are contracted. This propels the bolus in an aboral direction.
Peristalsis
Peristalsis is a sequential contraction of muscles in the GI system in response to stretching of the gut wall. The lumen is constricted immediately behind the bolus by the contraction of circular muscles, and there is relaxation of longitudinal muscles. Immediately in front of the bolus, there is relaxation of the circular muscles and contraction of the longitudinal muscles (Bayliss and Star-ling’s law of the intestines). This creates a pressure gradient that forces the bolus to move distally.
This coordinated motor activity is called the peristaltic reflex and is mediated by the enteric nervous system (Fig. 20.2).
20.1 Chewing and Swallowing
Chewing
Food enters the GI system via the mouth, where mastication (chewing) begins the process of mechanical breakdown.
Saliva lubricates the food and contains some digestive enzymes, for example, salivary lipase (very minor action) and salivary amylase.
The chewed food mixes with saliva to form a bolus. The tongue propels the bolus of food to the back of the oral cavity, initiating a swallowing reflex.
Muscles of mastication
The muscles of mastication are the masseter, the temporalis, and the medial and lateral pterygoids. They are innervated by the mandibular division of the trigeminal nerve (cranial nerve [CN] V).
Swallowing
Swallowing Reflex
The initial phase of swallowing is voluntary; the later phases are involuntary. Pressure receptors in the pharynx are stimulated by the presence of the bolus to send afferent signals, via the vagus and glossopharyngeal nerves, to the swallowing center in the medulla. Efferent signals are sent back from the medulla to striated muscles in the pharynx, larynx, and upper esophagus and to smooth muscles in the lower esophagus and stomach, coordinating a sequence of involuntary events that bring food from the mouth to the stomach via the pharynx and esophagus.
Pharyngeal phase of swallowing. The act of the tongue pushing the bolus into the pharynx elevates the soft palate and uvula to seal off the nasal cavity. Respiration is momentarily inhibited, the larynx is raised, and the epiglottis seals over the glottis so that food cannot enter the trachea. Peristalsis of the striated muscles of the pharynx force the bolus through the pharynx toward the esophagus. At the same time, the upper esophageal sphincter (UES) relaxes, allowing the bolus to pass, then closes to prevent regurgitation. Pressure in the resting esophagus parallels intrapleural pressure and thus is subatmospheric during inspiration. Therefore, closure of the UES also prevents entry of air into the esophagus during inspiration.
Esophageal phase of swallowing. The esophagus is gated on both ends by sphincters.
– The UES is composed of striated muscle and is normally closed due to elasticity of the sphincter and tonic neural excitation.
– The lower esophageal sphincter (LES) is composed of smooth muscle. It is closed under resting conditions due in part to tonic myogenic contraction (i.e., the contraction is due to an intrinsic property of the myocyte cell, not due to neural stimulation). The LES relaxes at the start of the swallow, several seconds before the wave of peristalsis in the esophagus approaches the LES due to vagal (noncholinergic) inhibition, allowing the bolus to pass into the stomach. It then closes to prevent reflux of gastric contents into the esophagus.
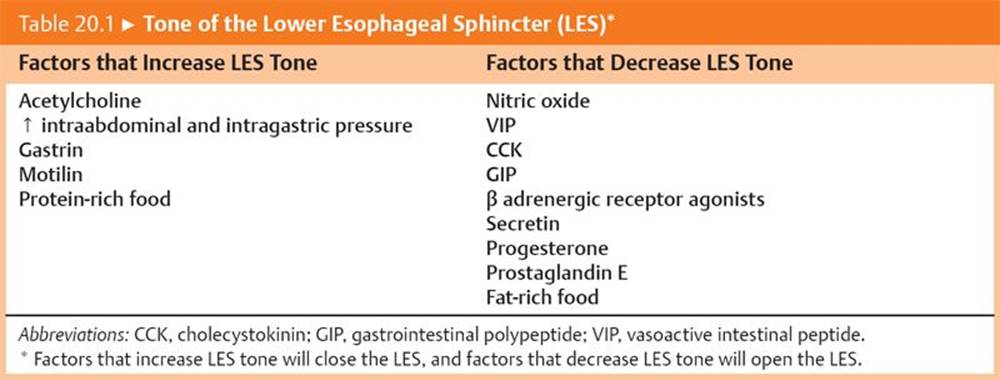
Table 20.1 lists the factors that modulate the tone of the LES.
Gastroesophageal reflex disease
Gastroesophageal reflux disease (GERD) occurs when stomach acid continuously refluxes into the esophagus. It may be caused by elevated intraabdominal pressure (e.g., due to obesity, big meals, or tight clothing), reduced LES tone (e.g., due to pregnancy, hiatus hernia, achalasia, fatty meals, and smoking), and by tricyclic and anticholinergic drugs. GERD causes pain, heartburn, and inflammation because the esophagus lacks the protective lining of the stomach. The pain of GERD radiates to the back and is worsened by stooping and ingesting hot drinks. Treatment is with antacids (e.g., calcium carbonate), H2-receptor antagonists (e.g., cimetidine), or proton pump inhibitors (e.g., omeprazole). Medication to strengthen the LES, known as prokinetic drugs (e.g., metoclopramide), may also be used. If medications alone do not control symptoms, surgery to tighten the LES may be necessary.
In an upright posture, swallowed liquids can simply flow down the esophagus due to the force of gravity (although gravity is not essential for swallowing). The passage of semisolid food down the esophagus requires peristalsis. Peristalsis in the esophagus is coordinated by the extrinsic and intrinsic nervous systems.
– Primary peristalsis automatically occurs when a bolus enters the esophagus. It is coordinated by extrinsic (autonomic nervous system) innervation of striated muscle.
– Secondary peristalsis is triggered by local distention of the esophagus. It moves any food left behind, and it returns material refluxed up from the stomach. It is coordinated by intrinsic (enteric nervous system) innervation of smooth muscle (Fig. 20.3).
Achalasia
Achalasia is a pathological condition caused by a deficiency of inhibitory myenteric neurons in the lower part of the esophagus. The LES fails to relax during swallowing, and peristalsis is absent. Food therefore accumulates above the LES, causing an increased risk of aspiration pneumonia. Symptoms of achalasia include regurgitation of food, chest pain, difficulty swallowing liquids and solids, cough, and weight loss. Drug treatment is aimed at reducing the tone of the LES. This may be achieved with Botox injections (temporary action) or by administration of long-acting nitrates or Ca2+ channel blockers. Surgical treatment includes esophagomyomectomy to reduce LES tone and dilation of the esophagus.
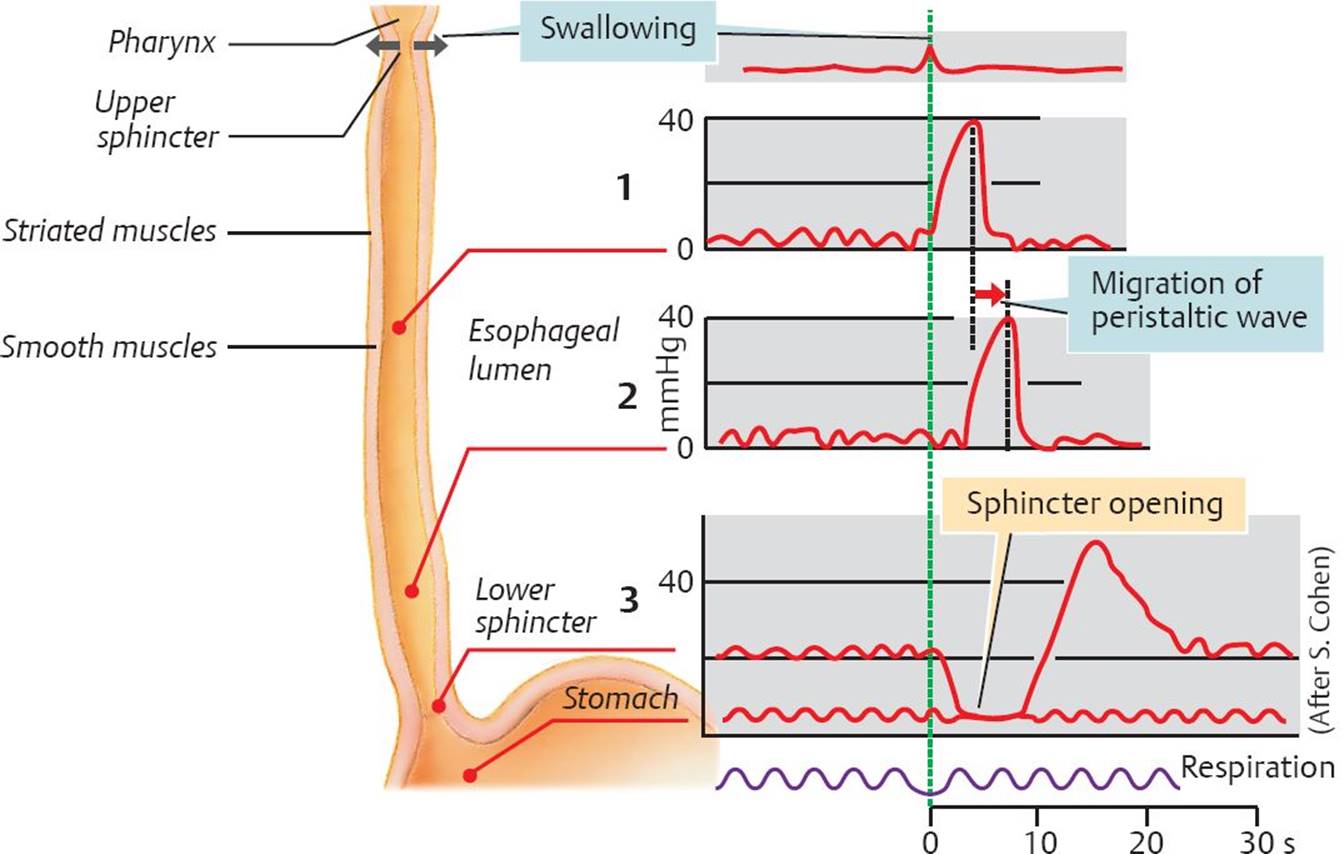
Fig. 20.3 ![]() Esophageal motility.
Esophageal motility.
Esophageal motility is usually checked by measuring the pressure in the lumen, for example, during a peristaltic wave (1,2). The resting pressure within the lower esophageal sphincter (LES) is normally 20 to 25 mm Hg. During receptive relaxation, esophageal pressure drops to match the low pressure in the proximal stomach (3), indicating opening of the LES. The LES opens due to a vagovagal reflex mediated by myenteric neurons releasing vasoactive intestinal peptide and nitric oxide.
20.2 Gastric Motility
In the stomach, chewed food is converted into a thick liquid called chyme by the addition of water and electrolytes (e.g., Na+, K+, and Ca2+). It is then mixed with acid and pepsin to begin the digestion process. Chyme is stored for variable amounts of time and then released in a slow, controlled fashion into the duodenum.
Functional Gastric Anatomy
– The proximal stomach is made up of the fundus and the proximal part of the corpus (body) (Fig. 20.4). The proximal stomach can easily expand to allow for increased volume of stomach contents.
– The distal stomach is made up of the distal part of the corpus, the antrum, and the pylorus (gastroduodenal junction [GDJ]), where the stomach and small intestine meet.
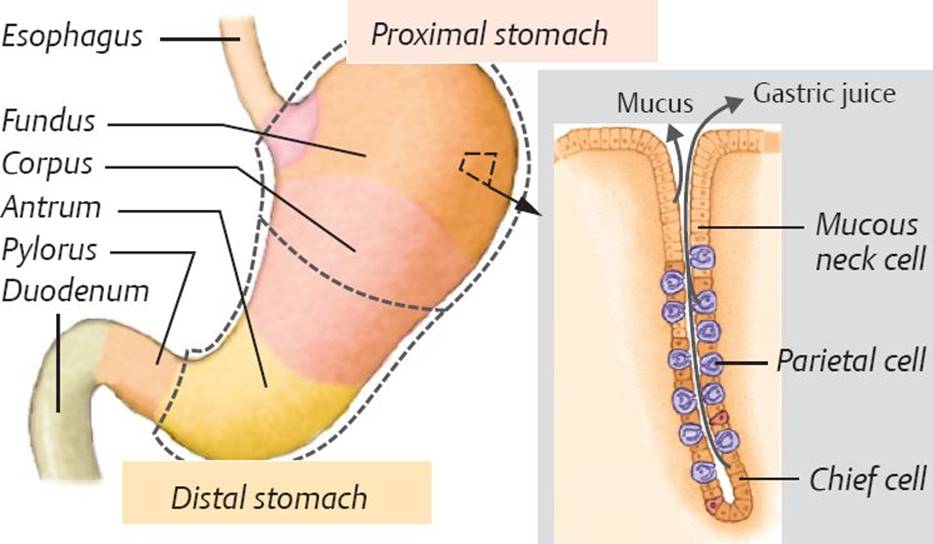
Fig. 20.4 ![]() Anatomy of the stomach.
Anatomy of the stomach.
The fundus of the stomach is continuous with the body (corpus), followed by the antrum. The pylorus connects the stomach to the duodenum. The mucosa of the fundus and corpus contain chief cells that produce pepsin, as well as parietal cells that produce gastric acid. The mucosa also contains endocrine cells that produce hormones (e.g., gastrin), mucous neck cells that produce protective mucus during feeding, and goblet cells that produce mucus continuously. The stomach is divided functionally into a proximal and distal segment. The proximal stomach mainly serves as a reservoir for food and propels gastric contents to the distal stomach, which is mainly concerned with mixing and pulverizing.
Receptive Relaxation
Receptive relaxation is a vagovagal reflex that causes the muscles of the proximal stomach to relax, which facilitates entry of the bolus into the stomach. It allows the stomach to expand without increasing intragastric pressure. This property is known as accommodation.
Receptive relaxation is mediated by vasoactive intestinal peptide (VIP).
Mixing and Digestion
Slow-wave depolarizations and peristaltic contractions usually arise in the corpus and spread distally, forcing chyme toward the pyloric sphincter at the GDJ (Fig. 20.5). Contractions are weak in the body of the stomach, where smooth muscle layers are relatively thin. Contractions are more forceful in the more muscular antrum. As the antrum begins to contract, some chyme may pass through the GDJ into the duodenal bulb. As antral contractions continue (antral systole), the pyloric sphincter closes, and the bulk of the chyme is retropulsed back into the body of the stomach; the process then repeats. Peristaltic contractions and antral systole function to mix gastric contents and to break up digestible solids, allowing them to be suspended in, and emptied along with, the liquid.
– Gastric peristalsis is stimulated by long and short cholinergic reflexes initiated by distention and by elevated serum gastrin. Elevated serum gastrin not only stimulates contractions but also increases the frequency of slow-wave depolarizations.
Migrating Myoelectric Complex
During fasting, the stomach exhibits regular contractile activity every 90 to 120 minutes called migrating myoelectric complex (MMC). The contractions begin slowly and increase in strength, culminating in powerful peristaltic contractions. The pyloric sphincter remains open, allowing the contractions to remove large (> 1 mm) nondigestible solids left behind in the stomach and small intestine. MMC also removes mucus, sloughed cells, and bacteria from the small intestine, helping to prevent bacterial overgrowth.
– MMC is controlled by motilin.
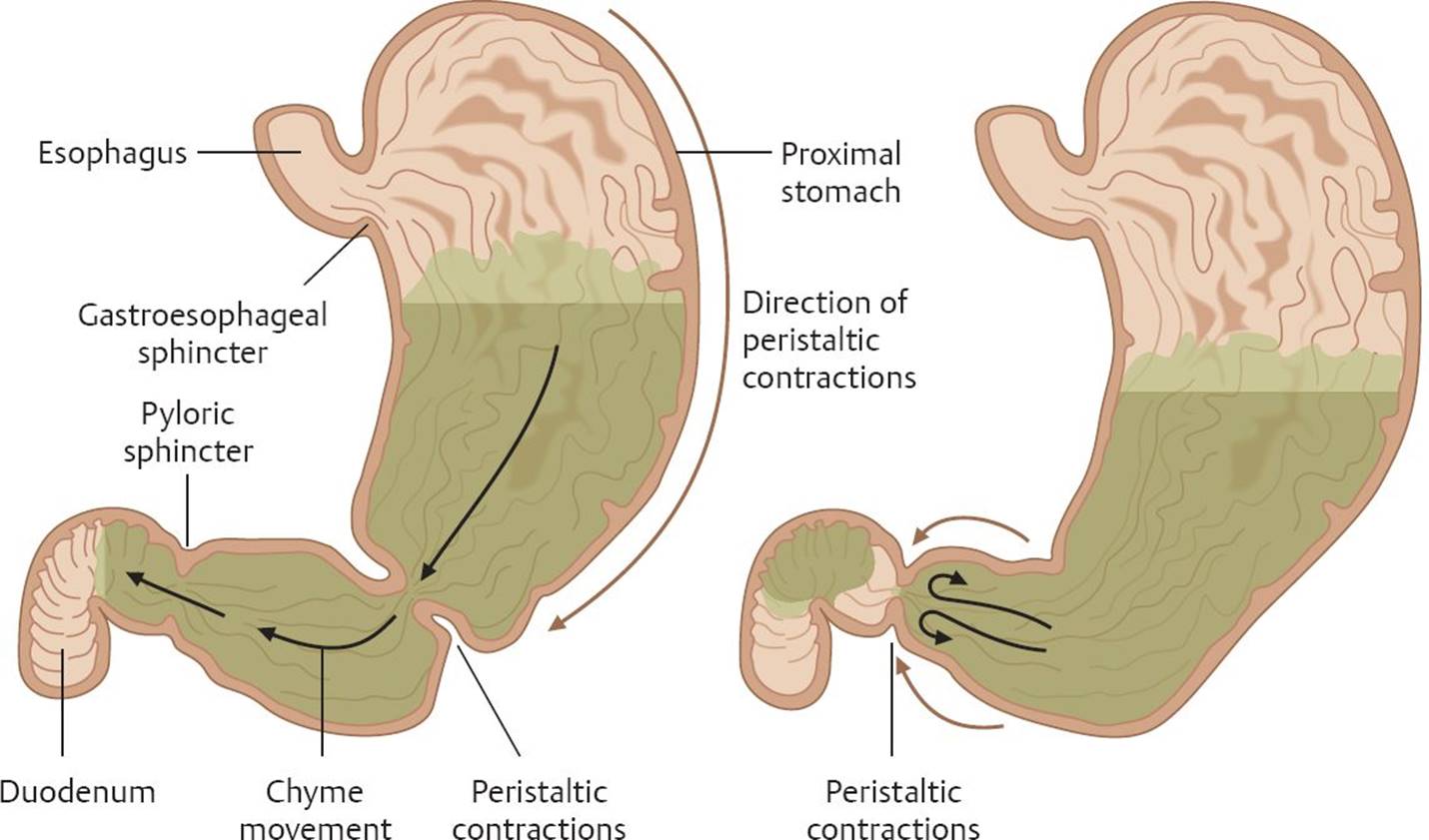
Fig. 20.5 ![]() Mixing in the distal stomach.
Mixing in the distal stomach.
Peristaltic contractions, originating in the corpus, drive stomach contents toward the pylorus. When the pyloric sphincter at the gastroduodenal junction closes, chyme is propelled back into the body of the stomach. This contributes to the breaking up of food particles, along with mixing and digestion of stomach contents.
Gastric Emptying
Chyme that leaves the stomach enters the duodenum as fluid and small solids (< 0.25 mm in diameter). While peristaltic contractions of the distal stomach aid in gastric emptying, elevation of intragastric pressure due to the sustained contractions of the proximal stomach contributes to emptying of gastric contents.
Rate of Gastric Emptying
– The rate of emptying is faster for liquids, small solids, and carbohydrates. It also increases with increased volume of chyme.
– The rate of emptying is slower for proteins and fats. It is also slower with increased acidity or osmolarity.
Control of Gastric Emptying: Enterogastric Reflex
Gastric emptying is principally regulated by the duodenum through the enterogastric reflex. The enterogastric reflex limits the amount of chyme that enters the duodenum at any one time so that digestion and absorption can proceed optimally. This reflex is mediated by hormonal and neural inputs.
Hormonal mediation of the enterogastric reflex
– Acidic chyme in the duodenum stimulates the release of secretin, which reduces gastric motility and increases the tone of the pyloric sphincter.
– The products of lipid digestion stimulate the release of cholecystokinin and glucose-dependent insulinotropic peptide (GIP), which also reduces gastric motility.
– The products of protein digestion stimulate the release of gastrin, cholecystokinin, and GIP, which all slow gastric emptying.
Neural mediation of the enterogastric reflex
– Distention of the duodenum activates duodenal stretch receptors that act via local neuronal circuits to slow gastric emptying.
20.3 Small Intestine Motility
The small intestine is the major site for digestion and absorption of nutrients from food. It receives 7 to 10 L of chyme per day from the stomach, to which various secretions are added.
The basic frequency and patterns of contractions in the small intestine are determined by slow-wave depolarizations. The frequency of slow-wave depolarization is highest in the proximal small intestine, ~12/min, and gradually decreases distally to become ~8/min in the terminal ileum. As in the stomach, the slow waves are myogenic and are not themselves sufficient to elicit contractions. Action potentials, superimposed upon the plateau of the slow wave, cause contraction of smooth muscles of the small intestine.
Entry into the Small Intestine
Contractions of the first section of the duodenum are coordinated with gastric contractions. When the pyloric sphincter is open, the bulb (opening) of the duodenum is relaxed, facilitating emptying of stomach contents and minimizing reflux of duodenal contents into the stomach. Following closure of the pyloric sphincter, the duodenal bulb contracts, moving the contents distally. When the small intestine is distended, stretch receptors are activated. Enteric neurons cause contractions in the circular muscle proximal to the distention and relaxation of longitudinal muscles. They also cause relaxation of the circular muscles and contraction of the longitudinal muscles distal to it. This activity is known as Bayliss and Starling’s law of the intestine.
– Overdistention of one portion of the small intestine results in inhibition of motility in the rest of the small intestine. This is known as the intestino-intestinal reflex.
Mixing of Chyme and Motility in the Small Intestine
Segmentation Contractions
– Segmentation contractions mix intestinal contents with digestive enzymes and bile salts. Contraction of an intestinal segment propels the chyme in a proximal and distal direction. Relaxation then causes the chyme to occupy the same segment as before; thus, there is no forward propulsion of chyme.
Peristaltic Contractions
– Peristaltic contractions are directed aborally (away from the mouth) and progressively move chyme through the small intestine.
– They can be initiated anywhere along the length of the small intestine and are propagated for distances of a few centimeters.
Regulation of Motility in the Small Intestine
Coordination of stomach contractions and contractions within the small intestine is mediated by the enteric nervous system and hormones.
– Small intestine motility is stimulated by parasympathetic neural activity, serotonin, gastrin, cholecystokinin, and motilin.
– Small intestine motility is inhibited by sympathetic neural activity, epinephrine, secretin, and glucagon.
Emptying of the Small Intestine
Gastroileal Reflex
Increased peristalsis in the ileum and relaxation of the ileocecal sphincter in response to the presence of food in the stomach is referred to as the gastroileal reflex. This reflex results in chyme leaving the small intestine and entering the large intestine.
– The gastroileal reflex is mediated by increased gastrin levels or through extrinsic neural reflexes.
20.4 Vomiting
Vomiting is a mechanism for the rapid evacuation of gastric and duodenal contents. It is often preceded by nausea and increased salivation (water brash).
– The vomiting center in the medulla receives stimuli from visual pathways, motion sensors (CN VIII), vagal afferents from the GI tract, and from the chemoreceptive trigger zone in the fourth ventricle (Fig. 20.6).
Vomiting Reflex
The vomiting reflex starts with a single retrograde peristaltic contraction beginning in the middle of the small intestine that propels intestinal contents through a relaxed GDJ into the stomach. Inspiration occurs against a closed glottis, lowering intraesophageal pressure. The duodenum and antrum contract to prevent movement of chyme back into the small intestine. The abdominal muscles then forcibly contract (Valsalva maneuver), increasing intraabdominal pressure, which creates more pressure in the stomach than in the esophagus. This forces gastric contents into the esophagus. The larynx and hyoid bone are drawn forward, decreasing the tone of the UES and leading to the gastric and esophageal contents being expelled via the oral cavity.
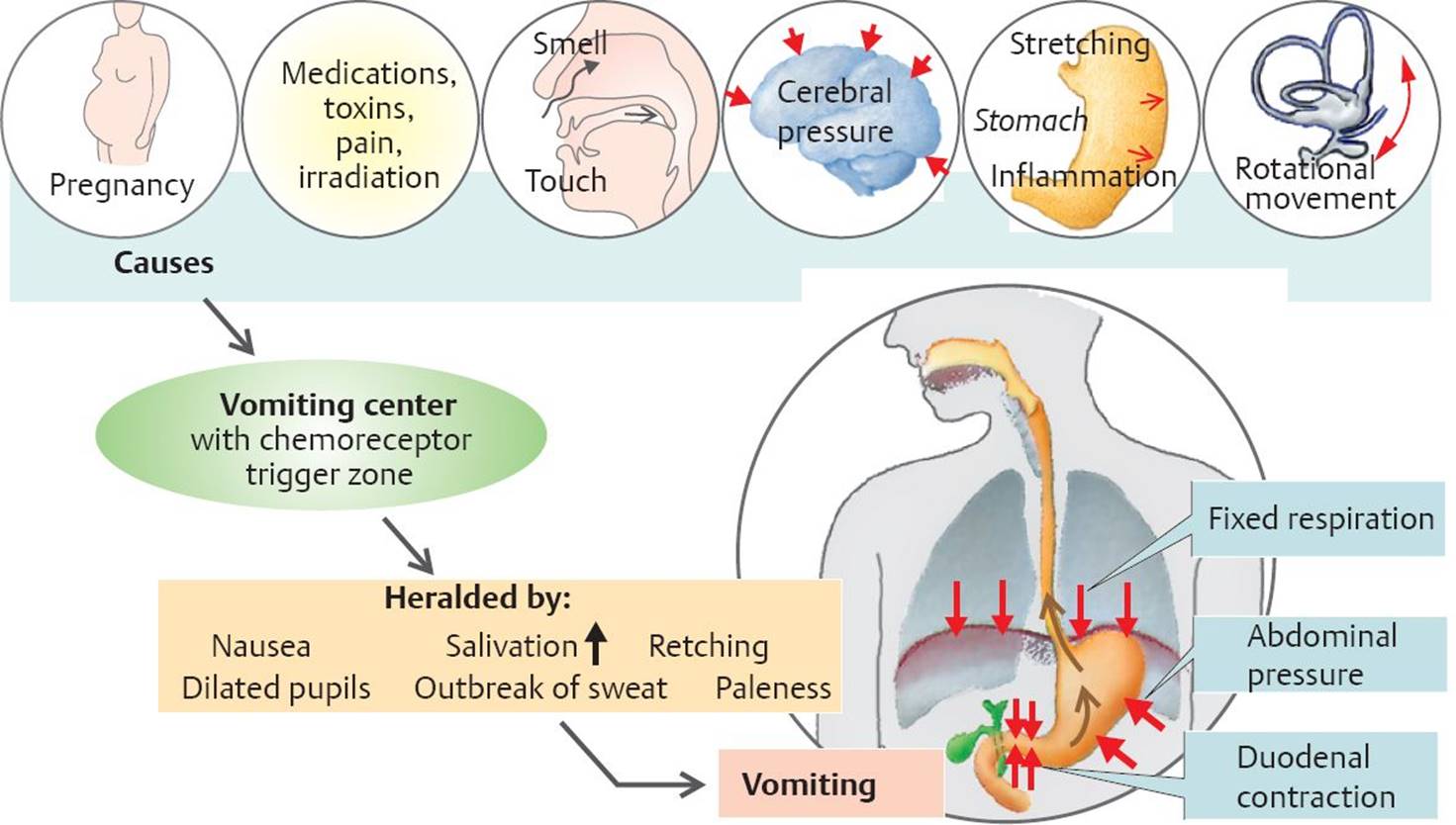
Fig. 20.6 ![]() Vomiting.
Vomiting.
Certain stimuli activate the vomiting center in the medulla oblongata, which receives input from chemosensors in the chemoreceptor trigger zone in the fourth ventricle. Vomiting is preceded by retching and increased salivation. During the act of vomiting, the diaphragm remains in the inspiratory position. The duodenum contracts, pushing intestinal contents into the stomach. The abdominal muscles contract, which then forces stomach contents into the esophagus. Relaxation of the upper esophageal sphincter results in ejection of stomach contents through the mouth.
20.5 Large Intestine Motility
The colon receives 0.5 to 1.0 L of chyme per day. Chyme moves very slowly from the cecum through the ascending, transverse, descending, and sigmoid parts of the colon. The arrangement of pacemakers in the colon means that retrograde movement is frequently seen, in contrast to the small intestine. This partly explains why colon transit time is longer than small intestine transit time. Chyme is mixed by low-frequency, long-duration contractions called haustrations, and fecal material is propelled through the colon by peristalsis.
In the colon, the predominant function of the enteric nerve plexus is to inhibit the innate desire of colonic smooth muscle to contract. The effect of removing this inhibition is highlighted in Hirschsprung disease.
Hirschsprung disease
Hirschsprung disease is a congenital absence of the enteric nervous system in a segment of the colon resulting in tonic c onstriction of the affected segment and dilation of the colon proximal to the constriction (megacolon). Symptoms in neonates include failure to pass stool within the first 2 days of life, vomiting, constipation or diarrhea, and gas. Treatment for this condition is surgery to remove the affected portion of bowel and connection of the remaining colon to the rectum. Sometimes an intervening colostomy is performed if the child is too ill to tolerate the full procedure.
Mass Movement
Periodically, haustrations cease, and colonic contents move distally due to large-scale peristaltic contractions. This mass movement occurs one to three times daily usually during or shortly after ingestion of the first meal of the day. This is most evident in infants. Stimulation of mass movement upon ingestion of a meal is called the gastrocolic reflex and may be triggered by increased gastrin or by extrinsic neural reflexes.
Defecation
The rectum normally contains little fecal material and is contracted to prevent entry of material from the colon. Occasionally, after a meal, mass movement shifts some of the colonic contents into the rectum. The resultant distention elicits the rectosphincter reflex that relaxes the internal anal sphincter and elicits the urge to defecate. If conditions are not appropriate for defecation, the external anal sphincter is voluntarily contracted, preventing expulsion of fecal material. When defecation is prevented, the rectum slowly pushes the material back into the sigmoid colon, and the internal anal sphincter regains its tone. Thus, the urge to defecate may be suppressed until the next mass movement causes the movement of fecal material from the sigmoid colon back into the rectum.
The act of defecation is partly involuntary and partly voluntary:
– Involuntary responses include contraction of smooth muscles of the distal colon and relaxation of the internal anal sphincter, mediated by the enteric nervous system.
– Voluntary movements include relaxation of the external anal sphincter (which is skeletal muscle that is innervated by the somatic nervous system) and voluntary contraction of abdominal muscles (Valsalva maneuver) to increase intraabdominal pressure to move feces out of the body.
Table 20.2 summarizes the GI reflexes that affect motility.