Input from the somatosensory systems informs the organism about events impinging on it. Sensation can be divided into four types: superficial, deep, visceral, and special. Superficial sensation is concerned with touch, pain, temperature, and two-point discrimination. Deep sensation includes muscle and joint position sense (proprioception), deep muscle pain, and vibration sense. Visceral sensations are relayed by autonomic afferent fibers and include hunger, nausea, and visceral pain (see Chapter 20). The special senses—smell, vision, hearing, taste, and equilibrium—are conveyed by cranial nerves (see Chapters 8, 15, 16, and 17). In addition, nociceptive sensation or pain-signaling serves to warn the organism when there is contact with noxious or potentially damaging elements in the environment, or when tissue is damaged.
RECEPTORS
Receptors are specialized cells for detecting particular changes in the environment. Exteroceptors include receptors affected mainly by the external environment: Meissner’s corpuscles, Merkel’s corpuscles, and hair cells for touch; Krause’s end-bulbs for cold; Ruffini’s corpuscles for warmth; and free nerve endings for pain (Fig 14–1). Receptors are not absolutely specific for a given sensation; strong stimuli can cause various sensations, even pain, even though the inciting stimuli are not necessarily painful. Proprioceptors receive impulses mainly from pacinian corpuscles, joint receptors, muscle spindles, and Golgi tendon organs. Painful stimuli are detected at the free endings of nerve fibers.
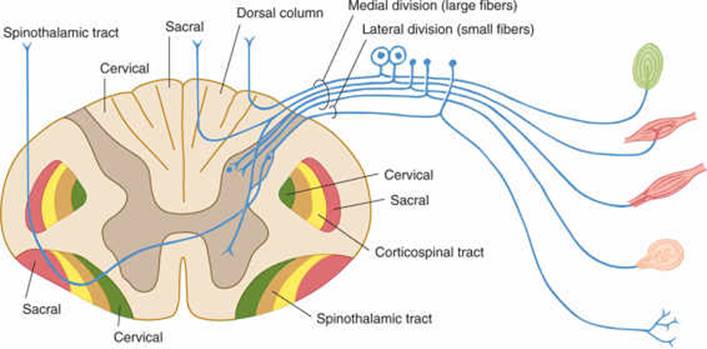
FIGURE 14–1 Schematic illustration of a spinal cord segment with its dorsal root, ganglion cells, and sensory organs.
Each efferent fiber from a receptor relays stimuli that originate in a receptive field and gives rise to a component of an afferent sensory system. Each individual receptor fires either completely or not at all when stimulated. The greater the intensity of a stimulus, the more end-organs that are stimulated, the higher the rate of discharge is, and the longer the duration of effect is. Adaptation denotes the diminution in rate of discharge of some receptors on repeated or continuous stimulation of constant intensity; the sensation of sitting in a chair or walking on even ground is suppressed.
CONNECTIONS
A chain of three long neurons and a number of interneurons conducts stimuli from the receptor or free ending to the somatosensory cortex (Figs 14–1 to 14–3).
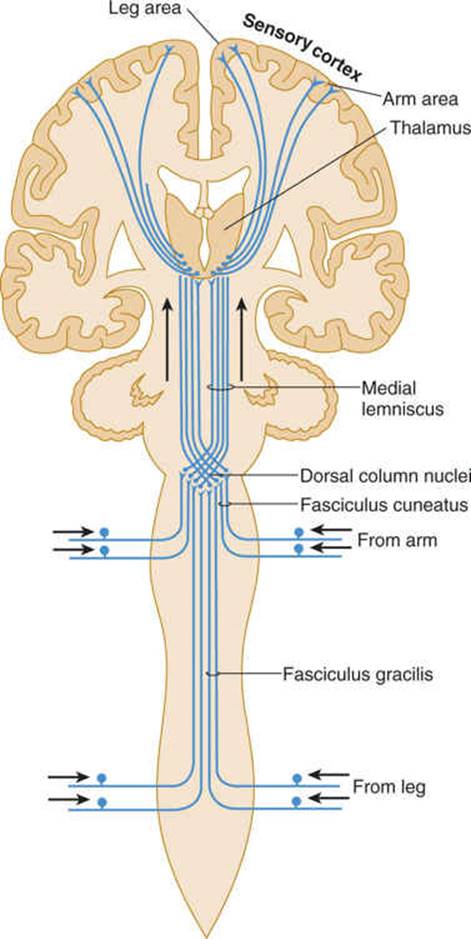
FIGURE 14–2 Dorsal column system for discriminative touch and position sense (lemniscus system).
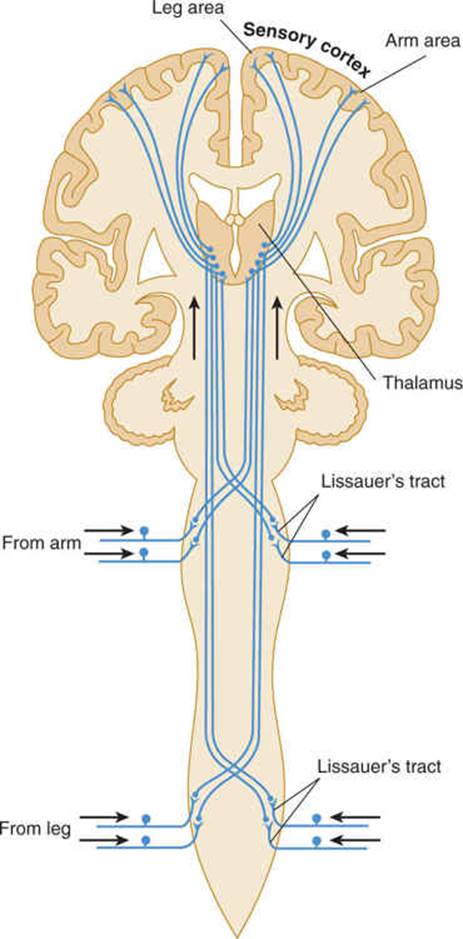
FIGURE 14–3 Spinothalamic tracts for pain and temperature (ventrolateral system).
First-Order Neuron
The cell body of a first-order neuron lies in a dorsal root ganglion or a somatic afferent ganglion (eg, trigeminal ganglion) of cranial nerves.
Second-Order Neuron
The cell body of a second-order neuron lies within the neuraxis (spinal cord or brain stem; examples are provided by the dorsal column nuclei, ie, the gracile and cuneate nuclei, and by neurons within the dorsal horn of the spinal cord). Axons of these cells usually decussate and terminate in the thalamus.
Third-Order Neuron
The cell body of a third-order neuron, which lies in the thalamus, projects rostrally to the sensory cortex. The networks of neurons within the cortex, in turn, process information relayed by this type of neuron; they interpret its location, quality, and intensity and make appropriate responses.
SENSORY PATHWAYS
Multiple neurons from the same type of receptor often form a bundle (tract), creating a sensory pathway. Sensory pathways ascending in the spinal cord are described in Chapter 5; their continuation within the brain stem is discussed in Chapter 7. The main sensory areas in the cortex are described in Chapter 10.
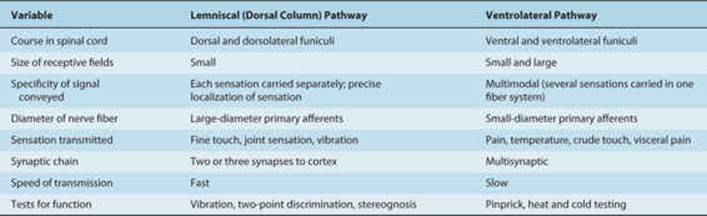
The lemniscal (dorsal column) system (see Fig 14–2) carries touch, joint sensation, two-point discrimination, and vibratory sense from receptors to the cortex. Another important system—the ventrolateral system—relays impulses concerning nociceptive stimuli (pain, crude touch) or changes in skin temperature (see Fig 14–3). Significant anatomic and functional differences characterize these two pathways: the size of the receptive field, nerve fiber diameter, course in the spinal cord, and function (Table 14–1). Each system is characterized by somatotopic distribution, with convergence in the thalamus (ventroposterior complex) and cerebral cortex (the sensory projection areas; see Figs 10–13 and 10–15), where there is a map-like representation of the body surface. The sensory trigeminal fibers contribute to both the lemniscal and the ventrolateral systems and provide the input from the face and mucosal membranes (see Figs 7–8 and 8–11).
TABLE 14–1 Differences between Lemniscal and Ventrolateral Systems.
CORTICAL AREAS
The primary somatosensory cortex (areas 3, 1, and 2) is organized in functional somatotopic columns that represent points in the receptive field. Within each column are inputs from thalamic, commissural, and associational fibers, all of which end in layers IV, III, and II (see Fig 10–10). The output is from cells in layers V and VI; however, the details of the processing occurring in each column and its functional significance (how it is felt) are largely unknown.
Additional cortical areas—secondary projection areas—also receive input from receptive fields in the columns. The somatotopic maps in these areas are more diffuse, however.
PAIN
Pathways
The free nerve endings that emanate from peripheral and cranial nerves are receptors, or nociceptors, for pain (see Figs 14–1 and 14–3). Nociceptors are sensitive to mechanical, thermal, or chemical stimuli. (Polymodal nociceptors are sensitive to several of these types of stimuli.) The pain fibers in peripheral nerves are of small diameter and are readily affected by local anesthetic. The thinly myelinated A-delta fibers convey discrete, sharp, short-lasting pain. The unmyelinated C fibers transmit chronic, burning pain. These nociceptive axons arise from small neurons located within the dorsal root ganglia and trigeminal ganglia.
Cells within injured tissue may release inflammatory molecules such as prostaglandins or other neuroactive molecules (such as histamine, serotonin, and bradykinin, in the aggregate comprising an “inflammatory soup”), which lower the threshold of peripheral nociceptors and thereby increase the sensibility to pain (hyperalgesia). Aspirin and other nonsteroidal anti-inflammatory drugs inhibit the action of prostaglandins and act to relieve pain (hypalgesiaor analgesia).
Pain Systems
The central projections of nociceptive primary sensory neurons impinge on second-order neurons within superficial layers of the dorsal horns of the spinal cord. According to the gate theory of pain, the strength of synaptic transmission at these junctions is decreased (probably by presynaptic inhibition) when large (non-pain-signaling) axons within the nerve are excited (the gate “closes”). Conversely, the strength of synaptic transmission along the pain-signaling pathway is increased when there is no large-fiber input.
CLINICAL CORRELATIONS
Interruption in the course of first- and second-order neurons produces characteristic sensory deficits, which can be especially apparent when they involve sensitive areas such as the face or fingertips. An example is provided by sensory loss in the territory innervated by a particular nerve or spinal root when this nerve or root is injured.
Thalamic lesions may be characterized by loss of the ability to discriminate or localize simple crude sensations or by severe, poorly localized pain (thalamic pain).
Following nerve injury, dorsal root ganglion neurons, including nociceptors, turn off certain genes and turn on others. As a result, they produce a type of sodium channel that is not normally present within them, and this can result in spontaneous firing (even when a noxious stimulus is not present) or hyper-responsiveness (firing at a pathologically increased, abnormally high frequency in response to peripheral stimulation). This abnormal hyperexcitability of dorsal root ganglion neurons contributes to neuropathic pain (pain associated with nerve injury).
Nociceptive dorsal root ganglion neurons can also become hyperexcitable, sending pain signals toward the brain even when a painful stimulus is not present, as a result of mutations of the sodium channels within them. In inherited erythromelalgia (the “man on fire” syndrome), for example, gain-of-function mutations of sodium channels within nociceptive dorsal root ganglion neurons lower the threshold for activation for these sodium channels (making it easier to turn them “on”) and keep them “on” longer once they are activated. As a result, the nociceptors become hyperexcitable, generating pain signals even in the absence of painful stimuli. Mutations of sodium channels also cause some painful peripheral neuropathies.
Because these disorders involve altered ion channel function, they are referred to as channelopathies.
There is also some evidence for long-lasting changes, which may underlie chronic pain syndromes, in the dorsal horn after nerve injury. For example, after injury to C fibers, these fibers may degenerate and vacate their synaptic target sites on superficial second-order neurons within the dorsal horn. Sprouting of larger primary afferent axons may cause nonnociceptive inputs to excite these superficial dorsal horn neurons (which normally do not signal pain). This central sensitization may contribute to allodynia (perception of an innocuous stimulus as painful) or hyperpathia (perception of a mildly unpleasant stimulus as very painful).
The central ascending pathway for sensation consists of two systems: the spinothalamic tract and the phylogenetically older spinoreticulothalamic system. The first pathway conducts the sensation of sharp, stabbing pain; the second conveys deep, poorly localized, burning pain. Both pathways are interrupted when the ventrolateral quadrant of the spinal cord is damaged by trauma or in surgery, such as a cordotomy, deliberately performed to relieve pain; contralateral loss of all pain sensation results below the lesion (Fig 14–4). These pathways project rostrally to a network of circuits termed as the pain matrix within the brain.
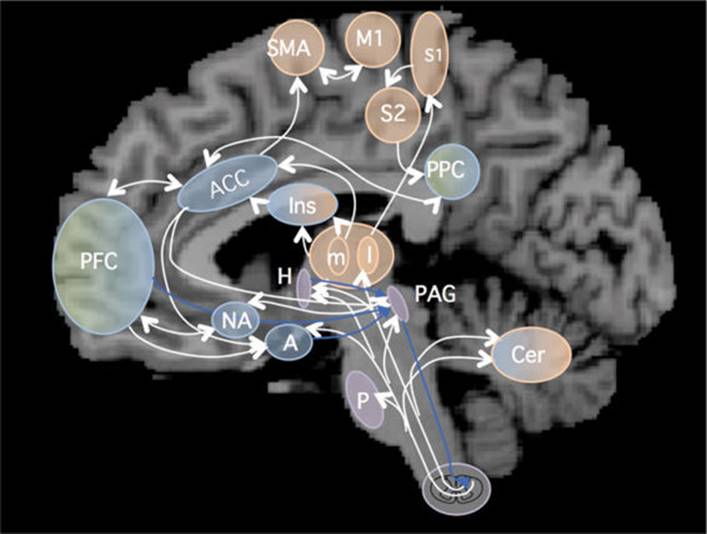
FIGURE 14–4 Overview of the pain matrix. White arrows: ascending and intracerebral pain pathways. Blue arrows: modulatory descending pathways. A, amygdale; ACC, anterior portion of cingulate cortex; Cer, cerebellum; H, hypothalamus; I, insula; L, m, lateral and medial thalamic nuclei; Mi, primary motor cortex; NA, nucleus accumbens; PAG, periaqueductal gray matter; PFC, prefrontal cortex; PPC, posterior parietal cortex; S1, S2, primary and secondary somatosensory cortex; SMA, supplementary motor area. (Reproduced, with permission, from Borsook D, Sava S, Becerra L: The pain imaging revolution: advancing pain into the 21st century, Neuroscientist 2009;16:172.)
The Pain Matrix
Pain elicits emotional and autononomic responses and is consciously appreciated as a result of activations of the pain matrix, which includes the thalamus, primary and secondary somatosensory cortex, insular cortex, prefrontal cortex, anterior cingulate cortex, supplementory motor area, posterior parietal cortex, periaqueductal gray matter, and amygdala, as well as the cerebellum (Fig 14–4).
Referred Pain
The cells in lamina V of the posterior column that receive noxious sensations from afferents in the skin also receive input from nociceptors in the viscera (Fig 14–5). When visceral afferents receive a strong stimulus, the cortex may misinterpret the source. A common example is referred pain in the shoulder caused by gallstone colic: The spinal segments that relay pain from the gallbladder also receive afferents from the shoulder region (convergence theory). Similarly, pain in the heart caused by myocardial infarct is conducted by fibers that reach the same spinal cord segments where pain afferents from the ulnar nerve (lower arm area) synapse.
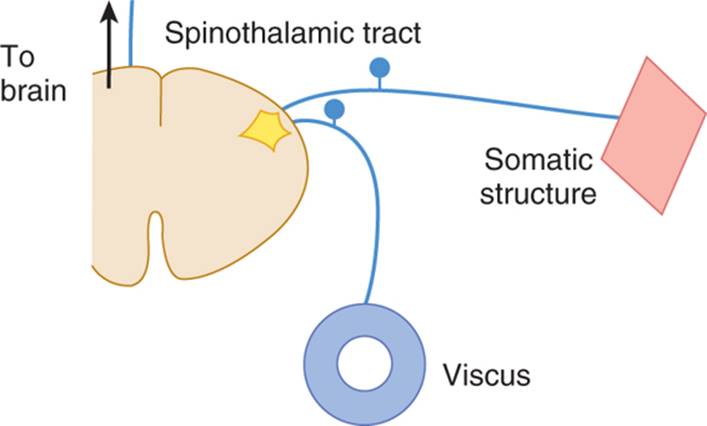
FIGURE 14–5 Diagram of convergence theory of referred pain. (Reproduced, with permission, from Ganong WF: Review of Medical Physiology. 22nd ed. McGraw-Hill, 2005.)
After injury to a peripheral nerve or root, some of the injured axons may generate inappropriate repetitive impulses, which can result in chronic pain. This is especially common when, as a result of an unsuccessful attempt at regeneration, sprouts from the injured axon form a tangle, or neuroma. After injury to these axons, dorsal root ganglion neurons can produce abnormal combinations of sodium and potassium channels, which cause them to generate inappropriate bursts of action potentials.
Descending Systems and Pain
Certain neurons within the brain, particularly within the periaqueductal gray matter of the midbrain, send descending axons to the spinal cord. These descending, inhibitory pathways suppress the transmission of pain signals and can be activated with endorphins and opiate drugs (Fig 14–6).
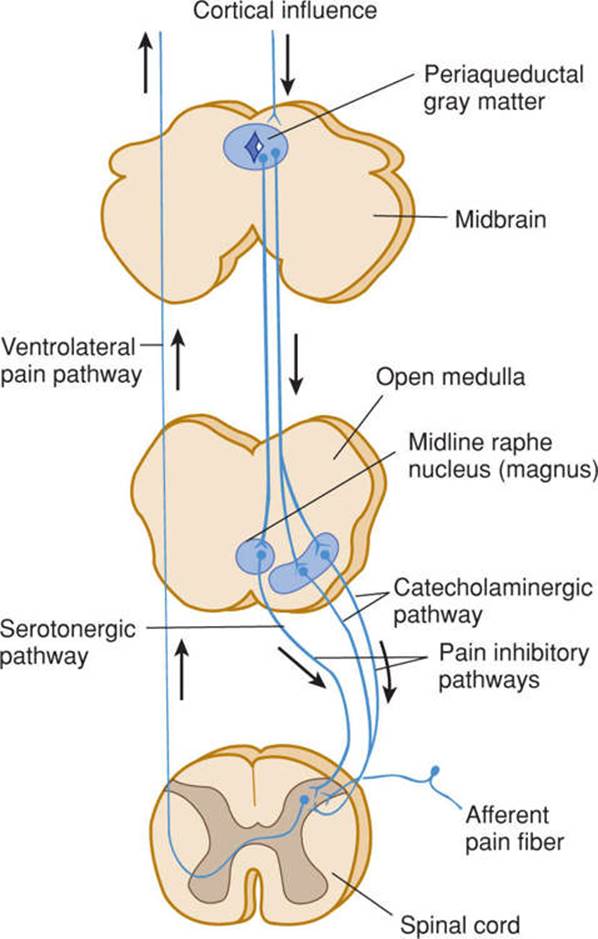
FIGURE 14–6 Schematic illustration of the pathways involved in pain control. (Used with permission from Al Basbaum.)
CASE 19
A 41-year-old woman complained of numbness and tingling in her right hand for more than a year. These sensations started gradually in the fingers but ultimately extended to the entire right hand and forearm. The patient was unable to do fine work such as sewing, and she sometimes dropped objects because of weakness that had developed in that hand. Three weeks before her admission to the hospital, she had burned two fingers of her right hand on her electric range; she had not felt the heat.
Neurologic examination showed wasting and weakness of the small muscles in the right hand. The deep tendon reflexes in the right upper extremity were absent or difficult to elicit. The knee and ankle jerks, however, were abnormally brisk, especially on the right side; the right plantar response was extensor. Abdominal reflexes were absent on both sides. Pain and temperature senses were lost in the right hand, forearm, and shoulder and in an area of the left shoulder. Touch, joint, and vibration senses were completely normal.
A plain-film radiograph of the spine was read as normal.
Where is the lesion? What is the differential diagnosis? Which imaging procedure(s) would be helpful? What is the most likely diagnosis?
CASE 20
A 41-year-old man was admitted to the hospital with progressive weakness and unsteadiness of his legs. His disability had begun more than a year earlier with tingling (“pins and needles”) feelings in his feet. Gradually, these sensations had become more disagreeable, and burning pains developed on the soles of his feet. The rest of his feet had become numb, and his legs had become weak. For about 6 months, he had had tingling feelings in his fingers and hands; his fingers felt clumsy, and he often dropped things. He had lost more than 6 kg (about 14 lbs) during the previous 6 months. The patient had smoked about 30 cigarettes daily for many years and drank eight glasses of beer and half a bottle of whiskey each day. After losing his job a year before, the patient had worked at several unskilled jobs.
Neurologic examination showed conspicuous atrophy in the calves, forearms, and intrinsic muscles of the hands. There was weakness of movement at both ankles and wrists and slightly weakened movement of the knees and elbows. The patient’s gait was unsteady and of the high-stepping type. There was loss of touch and pain sensation on the feet and distal thirds of the legs and on the hands and distal halves of the forearms, giving a “stocking-and-glove” distribution of sensory loss. Vibration sensibility was absent at the toes and ankles and diminished in the fingers. The soles of the feet and the calf muscles were hyperalgesic when squeezed. Ankle and biceps reflexes were absent, and knee jerks and triceps reflex were diminished.
What is the differential diagnosis? What is the most likely diagnosis?
Cases are discussed further in Chapter 25.
REFERENCES
Apkarian AV, Bushnell MC, Treede RD, Zubieta JK: Human brain mechanisms of pain perception and regulation in the health and disease. Europ J Pain 2005;9:463–484.
Borsook D, Sava S, Becerra L: The pain imaging revolution: Advancing pain into the 21st century. The Neuroscientist 2010;16:171–185.
Devor M, Rowbotham M, Wiessenfeld-Hallin Z: Progress in Pain Research and Management. IASP Press, 2000.
Dib-Hajj SD, Yang Y, Black JA, Waxman SG: The Nav1.7 sodium channel: from molecule to man. Nat Rev Neurosci. 2013;14:49–62.
Hoeijmakers JG, Faber CG, Lauria G, Merkies IS, Waxman SG: Small-fibre neuropathies: advances in diagnosis, pathophysiology and management. Nat Rev Neurol. 2012;8(7):369–379.
McMahon S, Koltzenberg M: Wall and Melzack’s Textbook of Pain, 5th ed. Elsevier, 2011.
Snyder WD, McMahon SB: Tackling pain at the source: New ideas about nociceptors. Neuron 1998;20:629.
BOX 14–1 Essentials for the Clinical Neuroanatomist
After reading and digesting this chapter, you should know and understand:
• First-, second-, and third-order neurons
• Dorsal column (lemniscal) pathway (Fig 14–2) and its function (Table 14–1)
• Ventrolateral pathways including spinothalamic tract (Fig 14–3) and their function (Table 14–1)
• Pain pathways and the pain matrix
• Neuroanatomic basis for referred pain