OBJECTIVES
After studying this chapter, you should be able to:
![]() Describe the primary types of rhythms that make up the electroencephalogram (EEG).
Describe the primary types of rhythms that make up the electroencephalogram (EEG).
![]() List the main clinical uses of the EEG.
List the main clinical uses of the EEG.
![]() Summarize the behavioral and EEG characteristics of each of the stages of nonrapid eye movement (NREM) and rapid eye movement (REM) sleep and the mechanisms responsible for their production.
Summarize the behavioral and EEG characteristics of each of the stages of nonrapid eye movement (NREM) and rapid eye movement (REM) sleep and the mechanisms responsible for their production.
![]() Describe the pattern of normal nighttime sleep in adults and the variations in this pattern from birth to old age.
Describe the pattern of normal nighttime sleep in adults and the variations in this pattern from birth to old age.
![]() Describe the interplay between brain stem neurons that contain norepinephrine, serotonin, and acetylcholine as well as GABA and histamine in mediating transitions between sleep and wakefulness.
Describe the interplay between brain stem neurons that contain norepinephrine, serotonin, and acetylcholine as well as GABA and histamine in mediating transitions between sleep and wakefulness.
![]() Discuss the circadian rhythm and the role of the suprachiasmatic nuclei (SCN) in its regulation.
Discuss the circadian rhythm and the role of the suprachiasmatic nuclei (SCN) in its regulation.
![]() Describe the diurnal regulation of synthesis of melatonin from serotonin in the pineal gland and its secretion into the bloodstream.
Describe the diurnal regulation of synthesis of melatonin from serotonin in the pineal gland and its secretion into the bloodstream.
INTRODUCTION
Most of the various sensory pathways described in Chapters 8–11 relay impulses from sense organs via three-and four-neuron chains to particular sites in the cerebral cortex. The impulses are responsible for perception and localization of individual sensations. However, they must be processed in the awake brain to be perceived. There is a spectrum of behavioral states ranging from deep sleep through light sleep, REM sleep, and the two awake states: relaxed awareness and awareness with concentrated attention. Discrete patterns of brain electrical activity correlate with each of these states. Feedback oscillations within the cerebral cortex and between the thalamus and the cortex serve as producers of this activity and possible determinants of the behavioral state. Arousal can be produced by sensory stimulation and by impulses ascending in the reticular core of the midbrain. Many of these activities have rhythmic fluctuations that are approximately 24 h in length; that is, they are circadian.
THALAMUS, CEREBRAL CORTEX, & RETICULAR FORMATION
THALAMIC NUCLEI
The thalamus is a large collection of neuronal groups within the diencephalon; it participates in sensory, motor, and limbic functions. Virtually all information that reaches the cortex is processed by the thalamus, leading to its being called the “gateway to the cerebral cortex.”
The thalamus can be divided into nuclei that project diffusely to wide regions of the neocortex and nuclei that project to specific discrete portions of the neocortex and limbic system. The nuclei that project to wide regions of the neocortex are the midline and intralaminar nuclei. The nuclei that project to specific areas include the specific sensory relay nuclei and the nuclei concerned with efferent control mechanisms. The specific sensory relay nucleiinclude the medial and lateral geniculate bodies, which relay auditory and visual impulses to the auditory and visual cortices; and the ventral posterior lateral (VPL) and ventral posteromedial nuclei, which relay somatosensory information to the postcentral gyrus. The ventral anterior and ventral lateral nuclei are concerned with motor function. They receive input from the basal ganglia and the cerebellum and project to the motor cortex. The anterior nuclei receive afferents from the mamillary bodies and project to the limbic cortex, which may be involved in memory and emotion. Most of the thalamic nuclei described are excitatory neurons that release glutamate. The thalamus also contains inhibitory neurons in the thalamic reticular nucleus. These neurons release GABA, and unlike the other thalamic neurons just described, their axons do not project to the cortex. Rather, they are thalamic interneurons and modulate the responses of other thalamic neurons to input coming from the cortex.
CORTICAL ORGANIZATION
The neocortex is generally arranged in six layers (Figure 14–1). The most common cell type is the pyramidal neuron with an extensive vertical dendritic tree (Figure 14–1 and Figure 14–2) that may reach to the cortical surface. Their cell bodies can be found in all cortical layers except layer I. The axons of these cells usually give off recurrent collaterals that turn back and synapse on the superficial portions of the dendritic trees. Afferents from the specific nuclei of the thalamus terminate primarily in cortical layer IV, whereas the nonspecific afferents are distributed to layers I–IV. Pyramidal neurons are the only projection neurons of the cortex, and they are excitatory neurons that release glutamate at their terminals. The other cortical cell types are local circuit neurons (interneurons) that have been classified based on their shape, pattern of projection, and neurotransmitter. Inhibitory interneurons (basket cellsand chandelier cells) release GABA as their neurotransmitter. Basket cells have long axonal endings that surround the soma of pyramidal neurons; they account for most inhibitory synapses on the pyramidal soma and dendrites. Chandelier cells are a powerful source of inhibition of pyramidal neurons because they have axonal endings that terminate exclusively on the initial segment of the pyramidal cell axon. Their terminal boutons form short vertical rows that resemble candlesticks, thus accounting for their name. Spiny stellate cells are excitatory interneurons that release glutamate as a neurotransmitter. These cells are located primarily in layer IV and are a major recipient of sensory information arising from the thalamus; they are an example of a multipolar neuron (Chapter 4) with local dendritic and axonal arborizations.
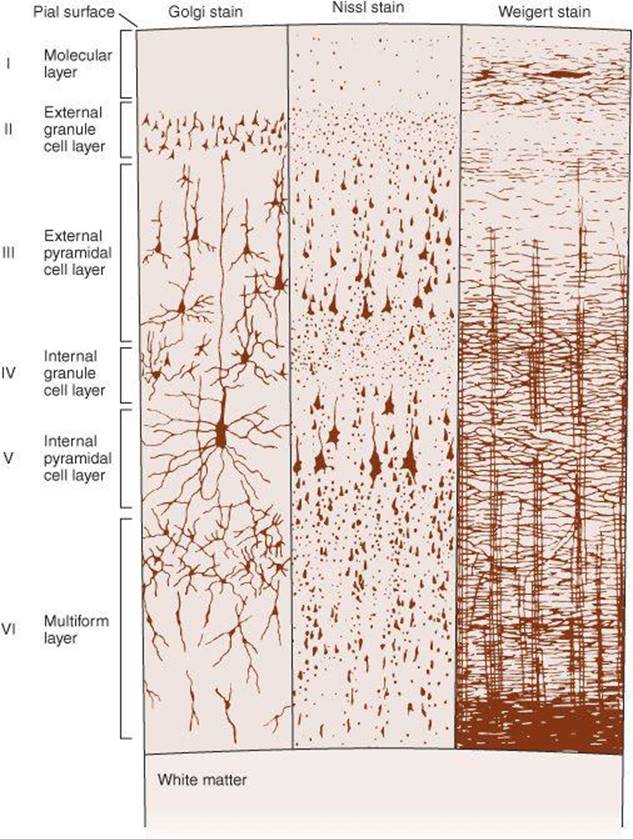
FIGURE 14–1 Structure of the cerebral cortex. The cortical layers are indicated by the numbers. Golgi stain shows neuronal cell bodies and dendrites, Nissl stain shows cell bodies, and Weigert myelin sheath stain shows myelinated nerve fibers. (Modified from Ranson SW, Clark SL: The Anatomy of the Nervous System, 10th ed. Saunders, 1959.)
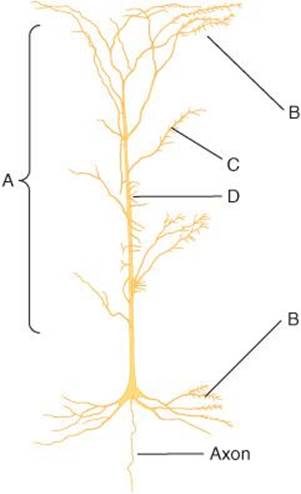
FIGURE 14–2 Neocortical pyramidal cell, showing the distribution of neurons that terminate on it. A denotes nonspecific afferents from the reticular formation and the thalamus; B denotes recurrent collaterals of pyramidal cell axons; C denotes commissural fibers from mirror image sites in the contralateral hemisphere; D denotes specific afferents from thalamic sensory relay nuclei. (Based on Scheibel ME, Scheibel AB: Structural organization of nonspecific thalamic nuclei and their projection toward cortex. Brain Res 1967 Sep;6(1):60–94.)
In addition to being organized into layers, the cerebral cortex is also organized into columns. Neurons within a column have similar response properties, suggesting they comprise a local processing network (eg, orientation and ocular dominance columns in the visual cortex).
RETICULAR ACTIVATING SYSTEM
The reticular formation, the phylogenetically old reticular core of the brain, occupies the central portion of the medulla and midbrain, surrounding the fourth ventricle and cerebral aqueduct. The reticular formation contains the cell bodies and fibers of many of the serotonergic, noradrenergic, and cholinergic systems. These pathways were shown in Figure 7–2. The reticular formation also contains many of the areas concerned with regulation of heart rate, blood pressure, and respiration. The reticular formation plays an important role in determining the level of arousal, thus it is called the ascending reticular activating system (RAS).
The RAS is a complex polysynaptic pathway arising from the brain stem reticular formation and hypothalamus with projections to the intralaminar and reticular nuclei of the thalamus which, in turn, project diffusely and nonspecifically to wide regions of the cortex including the frontal, parietal, temporal, and occipital cortices (Figure 14–3). Collaterals funnel into it not only from the long ascending sensory tracts but also from the trigeminal, auditory, visual, and olfactory systems. The complexity of the neuron net and the degree of convergence in it abolish modality specificity, and most reticular neurons are activated with equal facility by different sensory stimuli. The system is therefore nonspecific, whereas the classic sensory pathways are specific in that the fibers in them are activated by only one type of sensory stimulation.

FIGURE 14–3 Cross-section through the midline of the human brain showing the ascending reticular activating system in the brainstem with projections to the intralaminar nuclei of the thalamus and the output from the intralaminar nuclei to many parts of the cerebral cortex. Activation of these areas can be shown by positive emission tomography scans when subjects shift from a relaxed awake state to an attention-demanding task.
EVOKED CORTICAL POTENTIALS
The electrical events that occur in the cortex after stimulation of a sense organ can be monitored with a recording electrode. If the electrode is over the primary receiving area for a particular sense, a surface-positive wave appears with a latency of 5–12 ms. This is followed by a small negative wave, and then a larger, more prolonged positive deflection frequently occurs with a latency of 20–80 ms. The first positive–negative wave sequence is the primary evoked potential; the second is the diffuse secondary response.
The primary evoked potential is highly specific in its location and can be observed only where the pathways from a particular sense organ end. The positive–negative wave sequence recorded from the surface of the cortex occurs because the superficial cortical layers are positive relative to the initial negativity, then negative relative to the deep hyperpolarization. The surface-positive diffuse secondary response, unlike the primary response, is not highly localized. It appears at the same time over most of the cortex and is due to activity in projections from the midline and related thalamic nuclei.
PHYSIOLOGIC BASIS OF THE ELECTROENCEPHALOGRAM
The background electrical activity of the brain in unanesthetized animals was first described in the 19th century. Subsequently, it was analyzed in systematic fashion by the German psychiatrist Hans Berger, who introduced the term electroencephalogram (EEG) to denote the recording of the variations in brain potential. The EEG can be recorded with scalp electrodes through the unopened skull or with electrodes on or in the brain. The term electrocorticogram (ECoG) is used for the recording obtained with electrodes on the pial surface of the cortex.
The EEG recorded from the scalp is a measure of the summation of dendritic postsynaptic potentials rather than action potentials (Figure 14–4). The dendrites of the cortical neurons are a forest of similarly oriented, densely packed units in the superficial layers of the cerebral cortex (Figure 14–1). Propagated potentials can be generated in dendrites. In addition, recurrent axon collaterals end on dendrites in the superficial layers. As excitatory and inhibitory endings on the dendrites of each cell become active, current flows into and out of these current sinks and sources from the rest of the dendritic processes and the cell body. The cell body–dendrite relationship is therefore that of a constantly shifting dipole. Current flow in this dipole produces wave-like potential fluctuations in a volume conductor (Figure 14–4). When the sum of the dendritic activity is negative relative to the cell body, the neuron is depolarized and hyperexcitable; when it is positive, the neuron is hyperpolarized and less excitable.
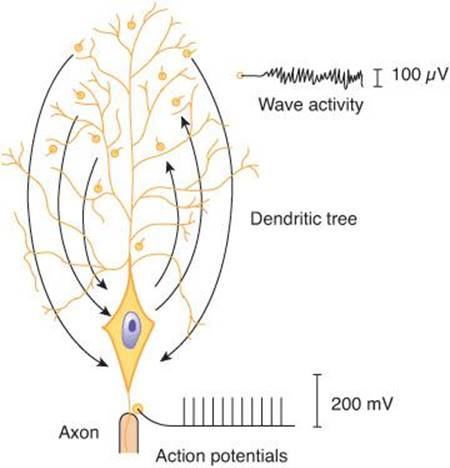
FIGURE 14–4 Diagrammatic comparison of the electrical responses of the axon and the dendrites of a large cortical neuron. Current flow to and from active synaptic knobs on the dendrites produces wave activity, while all-or-none action potentials are transmitted along the axon. When the sum of the dendritic activity is negative relative to the cell body, the neuron is depolarized; when it is positive, the neuron is hyperpolarized. The electroencephalogram recorded from the scalp is a measure of the summation of dendritic postsynaptic potentials rather than action potentials.
SLEEP–WAKE CYCLE: ALPHA, BETA, & GAMMA RHYTHMS
In adult humans who are awake but at rest with the mind wandering and the eyes closed, the most prominent component of the EEG is a fairly regular pattern of waves at a frequency of 8–13 Hz and amplitude of 50–100 μV when recorded from the scalp. This pattern is the alpha rhythm (Figure 14–5). It is most marked in the parietal and occipital lobes and is associated with decreased levels of attention. A similar rhythm has been observed in a wide variety of mammalian species. There are some minor variations from species to species, but in all mammals the pattern is remarkably similar (see Clinical Box 14–1).
CLINICAL BOX 14–1
Variations in the Alpha Rhythm
In humans, the frequency of the dominant EEG rhythm at rest varies with age. In infants, there is fast, beta-like activity, but the occipital rhythm is a slow 0.5–2-Hz pattern. During childhood this latter rhythm speeds up, and the adult alpha pattern gradually appears during adolescence. The frequency of the alpha rhythm is decreased by low blood glucose levels, low body temperature, low levels of adrenal glucocorticoid hormones, and high arterial partial pressure of CO2 (PaCO2). It is increased by the reverse conditions. Forced over-breathing to lower the PaCO2 is sometimes used clinically to bring out latent EEG abnormalities. The frequency and magnitude of the alpha rhythm is also decreased by metabolic and toxic encephalopathies including those due to hyponatremia and vitamin B12 deficiency. The frequency of the alpha rhythm is reduced during acute intoxication with alcohol, amphetamines, barbiturates, phenytoin, and antipsychotics. Propofol, a hypnotic/sedative drug, can induce a rhythm in the EEG that is analogous to the classic alpha rhythm.
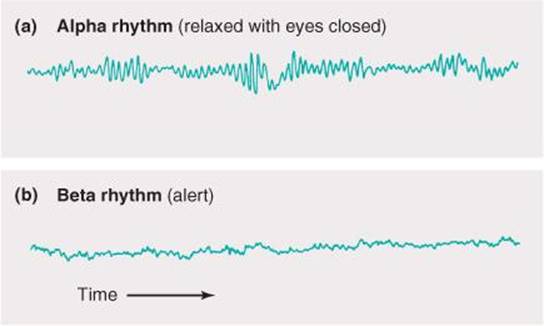
FIGURE 14–5 EEG records showing the alpha and beta rhythms. When attention is focused on something, the 8–13 Hz alpha rhythm is replaced by an irregular 13–30 Hz low-voltage activity, the beta rhythm. This phenomenon is referred to as alpha block, arousal, or the alerting response. (From Widmaier EP, Raff H, Strang KT: Vander’s Human Physiology, 11th ed. McGraw-Hill, 2008.)
When attention is focused on something, the alpha rhythm is replaced by an irregular 13–30 Hz low-voltage activity, the beta rhythm (Figure 14–5). This phenomenon is called alpha block and can be produced by any form of sensory stimulation or mental concentration, such as solving arithmetic problems. Another term for this phenomenon is the arousal or alerting response, because it is correlated with the aroused, alert state. It has also been called desynchronization, because it represents breaking up of the obviously synchronized neural activity necessary to produce regular waves. However, the rapid EEG activity seen in the alert state is also synchronized, but at a higher rate. Therefore, the term desynchronization is misleading. Gamma oscillations at 30–80 Hz are often seen when an individual is aroused and focuses attention on something. This is often replaced by irregular fast activity as the individual initiates motor activity in response to the stimulus.
THALAMOCORTICAL LOOP
A circuit linking the cortex and thalamus is thought to be important in generating patterns of brain activity in sleep–wake states. Figure 14–6 shows properties of activity in such a thalamocortical circuit hypothesized to be involved in generating rhythmic activity. Augmented activation of low threshold T-type Ca2+ channels in thalamic neurons likely contributes to both physiological and pathophysiological synchrony in thalamocortical circuits. Although not shown, inhibitory thalamic reticular neurons are elements of this network. The EEG shows the characteristic awake, light sleep, and deep sleep patterns of activity. Likewise, recordings from individual thalamic and cortical neurons show different patterns of rhythmic activity. In the waking state, corticocortical and thalamocortical networks generate higher-frequency rhythmic activity (30–80 Hz; gamma rhythm). This rhythm may be generated within the cells and networks of the cerebral cortex or within thalamocortical loops. The gamma rhythm has been suggested as a mechanism to “bind” together diverse sensory information into a single percept and action, but this theory is still controversial. In fact, disturbances in the integrity of this thalamocortical loop and its interaction with other brain structures may underlie some neurological disorders, including seizure activity.
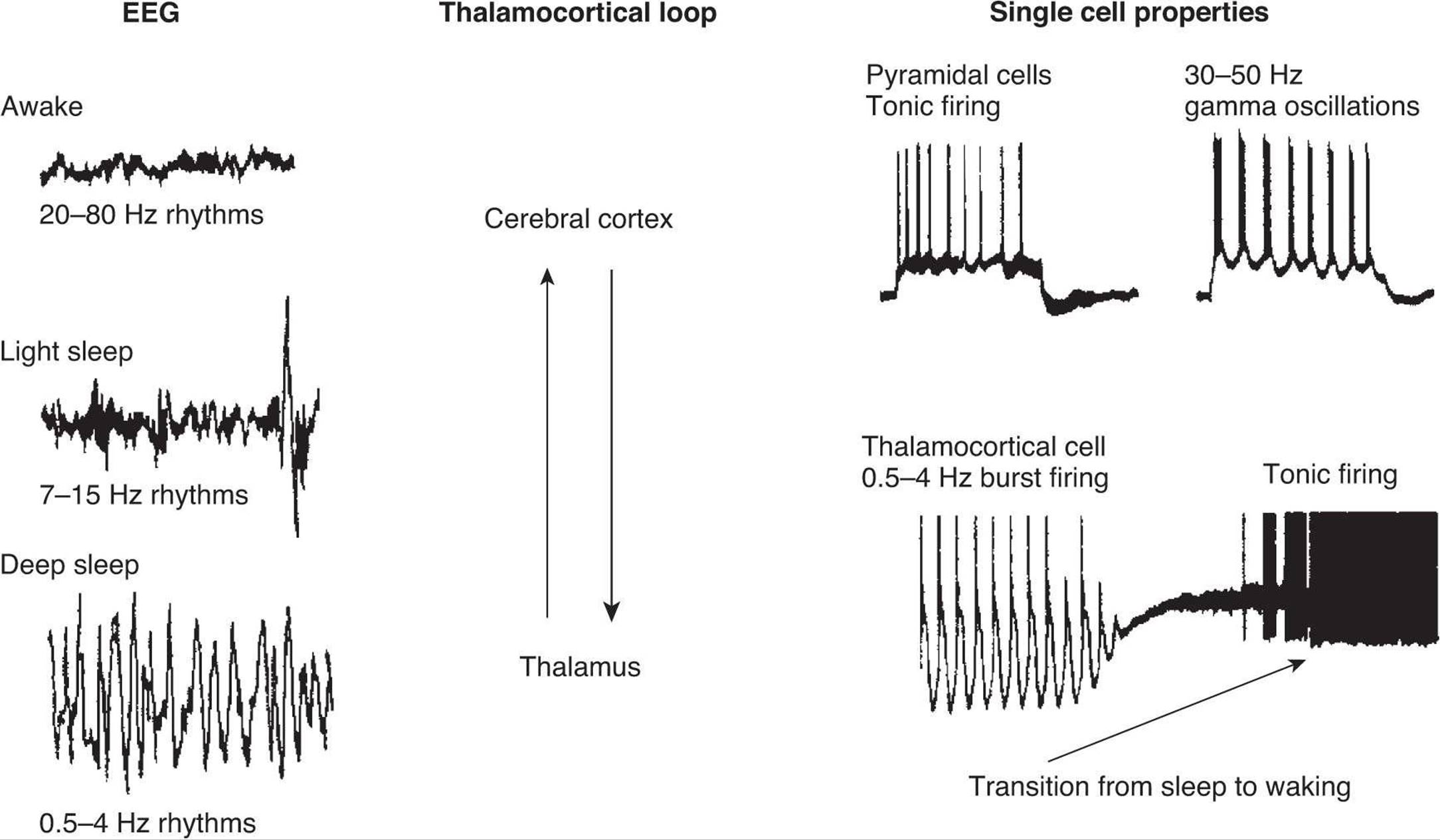
FIGURE 14–6 Correlation between behavioral states, EEG, and single-cell responses in the cerebral cortex and thalamus. The EEG is characterized by high-frequency oscillations in the awake state and low-frequency rhythms during sleep. Thalamic and cortical neurons can also show different patterns of rhythmic activity. Thalamocortical neurons show slow rhythmic oscillations during deep sleep, and fire tonic trains of action potentials in the awake state. Most pyramidal neurons in the cortex generate only tonic trains of action potentials, although others may participate in the generation of high frequency rhythms through activation of rhythmic bursts of spikes. The thalamus and cerebral cortex are connected together in a loop. (Modified from McCormick DA: Are thalamocortical rhythms the Rosetta stone of a subset of neurological disorders? Nat Med 1999;5:1349.)
SLEEP STAGES
There are two kinds of sleep: rapid eye movement (REM) sleep and nonREM (NREM), or slow-wave sleep. REM sleep is so named because of the characteristic eye movements that occur during this stage of sleep. NREM sleep is divided into four stages (Figure 14–7). As a person begins to fall asleep and enters stage 1, the EEG shows a low-voltage, mixed frequency pattern. A theta rhythm (4–7 Hz) can be seen at this early stage of slow-wave sleep. Throughout NREM sleep, there is some activity of skeletal muscle but no eye movements occur. Stage 2 of NREM sleep is marked by the appearance of sinusoidal waves called sleep spindles (12–14 Hz) and occasional high voltage biphasic waves called K complexes. In stage 3 of NREM sleep, a high-amplitude delta rhythm (0.5–4 Hz) dominates the EEG waves. Maximum slowing with large waves is seen in stage 4 of NREM sleep. Thus, the characteristic of deep sleep is a pattern of rhythmic slow waves, indicating marked synchronization; it is sometimes referred to as slow-wave sleep. While the occurrence of theta and delta rhythms is normal during sleep, their appearance during wakefulness is a sign of brain dysfunction.
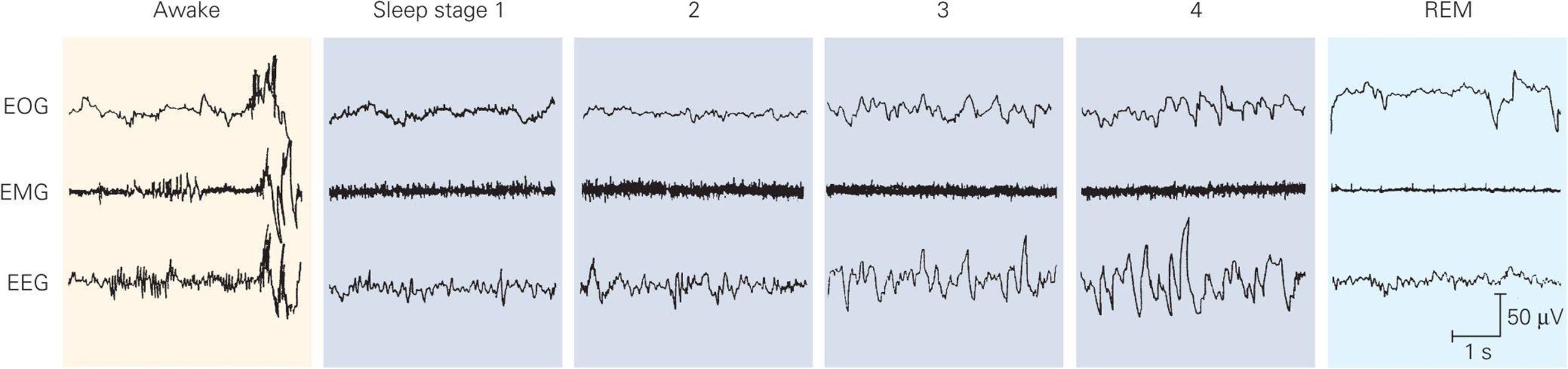
FIGURE 14–7 EEG and muscle activity during various stages of the sleep–wake cycle. NREM sleep has four stages. Stage 1 is characterized by a slight slowing of the EEG. Stage 2 has high-amplitude K complexes and spindles. Stages 3 and 4 have slow, high-amplitude delta waves. REM sleep is characterized by eye movements, loss of muscle tone, and a low-amplitude, high-frequency activity pattern. The higher voltage activity in the EOG tracings during stages 2 and 3 reflect high amplitude EEG activity in the prefrontal areas rather than eye movements. EOG, electro-oculogram registering eye movements; EMG, electromyogram registering skeletal muscle activity. (Reproduced with permission from Rechtschaffen A, Kales A: A Manual of Standardized Terminology, Techniques and Scoring System and Sleep Stages of Human Subjects. Los Angeles: University of California Brain Information Service, 1968.)
REM SLEEP
The high-amplitude slow waves seen in the EEG during sleep are periodically replaced by rapid, low-voltage EEG activity, which resembles that seen in the awake, aroused state and in stage 1 sleep (Figure 14–7). For this reason, REM sleep is also called paradoxical sleep. However, sleep is not interrupted; indeed, the threshold for arousal by sensory stimuli and by stimulation of the reticular formation is elevated. Rapid, roving movements of the eyes occur during paradoxical sleep, and it is for this reason that it is also called REM sleep. Another characteristic of REM sleep is the occurrence of large phasic potentials that originate in the cholinergic neurons in the pons and pass rapidly to the lateral geniculate body and from there to the occipital cortex. They are called pontogeniculo-occipital (PGO) spikes. The tone of the skeletal muscles in the neck is markedly reduced during REM sleep.
Humans aroused at a time when they show the EEG characteristics of REM sleep generally report that they were dreaming, whereas individuals awakened from slow-wave sleep do not. This observation and other evidence indicate that REM sleep and dreaming are closely associated.
Positron emission tomography (PET) scans of humans in REM sleep show increased activity in the pontine area, amygdala, and anterior cingulate gyrus, but decreased activity in the prefrontal and parietal cortex. Activity in visual association areas is increased, but there is a decrease in the primary visual cortex. This is consistent with increased emotion and operation of a closed neural system cut off from the areas that relate brain activity to the external world.
DISTRIBUTION OF SLEEP STAGES
In a typical night of sleep, a young adult first enters NREM sleep, passes through stages 1 and 2, and spends 70–100 min in stages 3 and 4. Sleep then lightens, and a REM period follows. This cycle is repeated at intervals of about 90 min throughout the night (Figure 14–8). The cycles are similar, though there is less stage 3 and 4 sleep and more REM sleep toward morning. Thus, 4–6 REM periods occur per night. REM sleep occupies 80% of total sleep time in premature infants and 50% in full-term neonates. Thereafter, the proportion of REM sleep falls rapidly and plateaus at about 25% until it falls to about 20% in the elderly. Children have more total sleep time (8–10 h) compared to most adults (about 6 h).
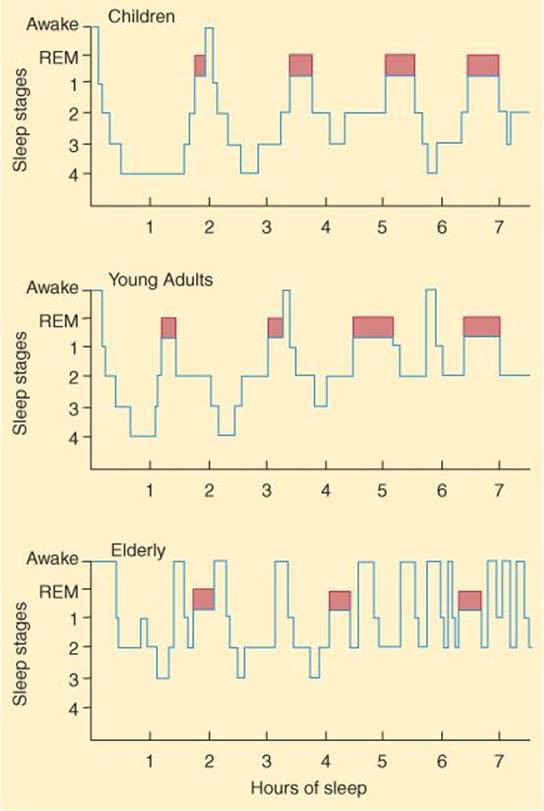
FIGURE 14–8 Normal sleep cycles at various ages. REM sleep is indicated by the darker colored areas. In a typical night of sleep, a young adult first enters NREM sleep, passes through stages 1 and 2, and spends 70–100 min in stages 3 and 4. Sleep then lightens, and a REM period follows. This cycle is repeated at intervals of about 90 min throughout the night. The cycles are similar, though there is less stage 3 and 4 sleep and more REM sleep toward morning. REM sleep occupies 50% of total sleep time in neonates; this proportion declines rapidly and plateaus at −25% until it falls further in the elderly. (Reproduced with permission from Kales AM, Kales JD: Sleep disorders. N Engl J Med 1974;290:487.)
IMPORTANCE OF SLEEP
Sleep has persisted throughout evolution of mammals and birds, so it is likely that it is functionally important. Indeed, if humans are awakened every time they show REM sleep, then permitted to sleep without interruption, they show a great deal more than the normal amount of REM sleep for a few nights. Relatively prolonged REM deprivation does not seem to have adverse psychological effects. However, rats deprived of all sleep for long periods lose weight in spite of increased caloric intake and eventually die. Various studies imply that sleep is needed to maintain metabolic-caloric balance, thermal equilibrium, and immune competence.
In experimental animals, sleep is necessary for learning and memory consolidation. Learning sessions do not improve performance until a period of slow-wave or slow-wave plus REM sleep has occurred. Clinical Box 14–2describes several common sleep disorders.
CLINICAL BOX 14–2
Sleep Disorders
Narcolepsy is a chronic neurological disorder caused by the brain’s inability to regulate sleep-wake cycles normally, and in which there is a sudden loss of voluntary muscle tone (cataplexy), an eventual irresistible urge to sleep during daytime, and possibly also brief episodes of total paralysis at the beginning or end of sleep. Narcolepsy is characterized by a sudden onset of REM sleep, unlike normal sleep that begins with NREM, slow-wave sleep. The prevalence of narcolepsy ranges from 1 in 600 in Japan to 1 in 500,000 in Israel, with 1 in 1000 Americans being affected. Narcolepsy has a familial incidence strongly associated with a class II antigen of the major histocompatibility complex on chromosome 6 at the HLA-DR2 or HLA-DQW1 locus, implying a genetic susceptibility to narcolepsy. The HLA complexes are interrelated genes that regulate the immune system (see Chapter 3). Compared to brains from healthy subjects, the brains of humans with narcolepsy often contain fewer hypocretin (orexin)-producing neurons in the hypothalamus. It is thought that the HLA complex may increase susceptibility to an immune attack on these neurons, leading to their degeneration.
Obstructive sleep apnea (OSA) is the most common cause of daytime sleepiness due to fragmented sleep at night and affects about 24% of middle-aged men and 9% of women in the United States. Breathing ceases for more than 10 s during frequent episodes of obstruction of the upper airway (especially the pharynx) due to reduction in muscle tone. The apnea causes brief arousals from sleep in order to reestablish upper airway tone. An individual with OSA typically begins to snore soon after falling asleep. The snoring gets progressively louder until it interrupted by an episode of apnea, which is then followed by a loud snort and gasp, as the individual tries to breathe. OSA is not associated with a reduction in total sleep time, but individuals with OSA experience a much greater time in stage 1 NREM sleep (from an average of 10% of total sleep to 30–50%) and a marked reduction in slow-wave sleep (stages 3 and 4 NREM sleep). The pathophysiology of OSA includes both a reduction in neuromuscular tone at the onset of sleep and a change in the central respiratory drive.
Periodic limb movement disorder (PLMD) is a stereotypical rhythmic extension of the big toe and dorsiflexion of the ankle and knee during sleep lasting for about 0.5–10 s and recurring at intervals of 20–90 s. Movements can actually range from shallow, continual movement of the ankle or toes, to wild and strenuous kicking and flailing of the legs and arms. Electromyograph (EMG) recordings show bursts of activity during the first hours of NREM sleep associated with brief EEG signs of arousal. The duration of stage 1 NREM sleep may be increased and that of stages 3 and 4 may be decreased compared to age-matched controls. PLMD is reported to occur in 5% of individuals between the ages of 30 and 50 years and increases to 44% of those over the age of 65. PLMD is similar to restless leg syndrome in which individuals have an irresistible urge to move their legs while at rest all day long.
Sleepwalking (somnambulism), bed-wetting (nocturnal enuresis), and night terrors are referred to as parasomnias, which are sleep disorders associated with arousal from NREM and REM sleep. Episodes of sleepwalking are more common in children than in adults and occur predominantly in males. They may last several minutes. Somnambulists walk with their eyes open and avoid obstacles, but when awakened they cannot recall the episodes.
THERAPEUTIC HIGHLIGHTS
Excessive daytime sleepiness in patients with narcolepsy can be treated with amphetamine-like stimulants, including modafinil, methylphenidate (Ritalin), and methamphetamine. Gamma hydroxybutyrate (GHB) is used to reduce the frequency of cataplexy attacks and the incidences of daytime sleepiness. Cataplexy is often treated with antidepressants such as imipramine and desipramine, but these drugs are not officially approved by the US Federal Drug Administration for such use. The most common treatment for OSA is continuous positive airflow pressure (CPAP), a machine that increases airway pressure to prevent airway collapse. Drugs have generally proven to have little or no benefit in treating OSA. Drugs used to treat Parkinson disease, dopamine agonists, can be used to treat PLMD.
CLINICAL USES OF THE EEG
The EEG is sometimes of value in localizing pathologic processes. When a collection of fluid overlies a portion of the cortex, activity over this area may be damped. This fact may aid in diagnosing and localizing conditions such as subdural hematomas. Lesions in the cerebral cortex cause local formation of transient disturbances in brain activity, marked by high-voltage abnormal waves that can be recorded with an EEG. Seizure activity can occur because of increased firing of neurons that are excitatory (eg, release of glutamate) or decreased firing of neurons that are inhibitory (eg, release GABA).
TYPES OF SEIZURES
Epilepsy is a condition in which there are recurring, unprovoked seizures that may result from damage to the brain. The seizures represent abnormal, highly synchronous neuronal activity. Epilepsy is a syndrome with multiple causes. In some forms, characteristic EEG patterns occur during seizures; between attacks; however, abnormalities are often difficult to demonstrate. Seizures are divided into partial (focal) seizures and generalized seizures.
Partial seizures originate in a small group of neurons and can result from head injury, brain infection, stroke, or tumor, but often the cause is unknown. Symptoms depend on the seizure focus. They are further subdivided into simple partial seizures (without loss of consciousness) and complex partial seizures (with altered consciousness). An example of a simple partial seizure is localized jerking movements in one hand progressing to clonic movements of the entire arm lasting about 60–90 s. Auras typically precede the onset of a partial seizure and include abnormal sensations. The time after the seizure until normal neurological function returns is called the postictal period.
Generalized seizures are associated with widespread electrical activity and involve both hemispheres simultaneously. They are further subdivided into convulsive and nonconvulsive categories depending on whether tonic or clonic movements occur. Absence seizures (formerly called petit mal seizures) are one of the forms of nonconvulsive generalized seizures characterized by a momentary loss of consciousness. They are associated with 3/s doublets, each consisting of a typical spike-and-wave pattern of activity that lasts for about 10 s (Figure 14–9). They are not accompanied by auras or postictal periods. These spike and waves are likely generated by low threshold T-type Ca2+channels in thalamic neurons.
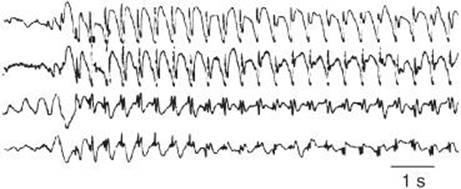
FIGURE 14–9 Absence seizures. This is a recording of four cortical EEG leads from a 6-year-old boy who, during the recording, had one of his “blank spells” in which he was transiently unaware of his surroundings and blinked his eyelids. Absence seizures are associated with 3/s doublets, each consisting of a typical spike-and-wave pattern of activity that lasts for about 10 s. Time is indicated by the horizontal calibration line. (Reproduced with permission from Waxman SG: Neuroanatomy with Clinical Correlations, 25th ed. McGraw-Hill, 2003.)
The most common convulsive generalized seizure is tonic-clonic seizure (formerly called grand mal seizure). This is associated with sudden onset of contraction of limb muscles (tonic phase) lasting about 30 s, followed by a clonic phase with symmetric jerking of the limbs as a result of alternating contraction and relaxation (clonic phase) lasting 1–2 min. There is fast EEG activity during the tonic phase. Slow waves, each preceded by a spike, occur at the time of each clonic jerk. For a while after the attack, slow waves are present.
Recent research provides insight into a possible role of release of glutamate from astrocytes in the pathophysiology of epilepsy. Also, there is evidence to support the view that reorganization of astrocytes along with dendritic sprouting and new synapse formation form the structural basis for recurrent excitation in the epileptic brain. Clinical Box 14–3 describes information regarding the role of genetic mutations in some forms of epilepsy.
CLINICAL BOX 14–3
Genetic Mutations & Epilepsy
Epilepsy has no geographical, racial, gender, or social bias. It can occur at any age, but is most often diagnosed in infancy, childhood, adolescence, and old age. It is the second most common neurological disorder after stroke. According to the World Health Organization, it is estimated that 50 million people worldwide (8.2 per 1000 individuals) experience epileptic seizures. The prevalence in developing countries (such as Colombia, Ecuador, India, Liberia, Nigeria, Panama, United Republic of Tanzania, and Venezuela) is more than 10 per 1000. Many affected individuals experience unprovoked seizures, for no apparent reason, and without any other neurological abnormalities. These are called idiopathic epilepsies and are assumed to be genetic in origin. Mutations in voltage-gated potassium, sodium, and chloride channels have been linked to some forms of idiopathic epilepsy. Mutated ion channels can lead to neuronal hyperexcitability via various pathogenic mechanisms. Scientists have recently identified the mutated gene responsible for development of childhood absence epilepsy (CAE). Several patients with CAE were found to have mutations in a subunit gene of the GABA receptor called GABRB3. Also, SCN1A and SCN1B mutations have been identified in an inherited form of epilepsy called generalized epilepsy with febrile seizures. SCN1A and SCN1B are sodium channel subunit genes that are widely expressed within the nervous system. SCN1A mutations are suspected in several other forms of epilepsy.
THERAPEUTIC HIGHLIGHTS
There are three broad mechanisms of action of new and old anticonvulsant drugs: enhancing inhibitory neurotransmission (increase GABA release), reducing excitatory neurotransmission (decrease glutamate release), or altering ionic conductance. Gabapentin is a GABA analog that acts by decreasing Ca2+ entry into cells and reducing glutamate release; it is used to treat generalized seizures. Topiramate blocks voltage-gated Na+ channels associated with glutamate receptors and potentiates the inhibitory effect of GABA; it is also used to treat generalized seizures. Ethosuximide reduces the low threshold T-type Ca2+ currents in thalamic neurons, and thus is particularly effective in treatment of absence seizures. Valproate and phenytoin block high frequency firing of neurons by acting on voltage-gated Na+ channels to reduce glutamate release.
TREATMENT OF SEIZURES
Only about 2/3 of those suffering from seizure activity respond to drug therapies. Some respond to surgical interventions (eg, those with temporal lobe seizures), whereas others respond to vagal nerve stimulation (eg, those with partial seizures). Prior to the 1990’s, the most common drugs used to treat seizures (anticonvulsants) included phenytoin, valproate, and barbiturates. Newer drugs have become available, but, as is the case with the older drugs, they are palliative rather than curative. Clinical Box 14–3 describes mechanisms of actions of some drugs used to treat seizures.
CIRCADIAN RHYTHMS & THE SLEEP–WAKE CYCLE
CIRCADIAN RHYTHMS
Most, if not all, living cells in plants and animals have rhythmic fluctuations in their function on a circadian cycle. Normally they become entrained, that is, synchronized to the day–night light cycle in the environment. If they are not entrained, they become progressively more out of phase with the light–dark cycle because they are longer or shorter than 24 h. The entrainment process in most cases is dependent on the suprachiasmatic nuclei (SCN) located bilaterally above the optic chiasm (Figure 14–10). These nuclei receive information about the light–dark cycle via a special neural pathway, the retinohypothalamic fibers. Efferents from the SCN initiate neural and humoral signals that entrain a wide variety of well-known circadian rhythms including the sleep–wake cycle and the secretion of the pineal hormone melatonin.
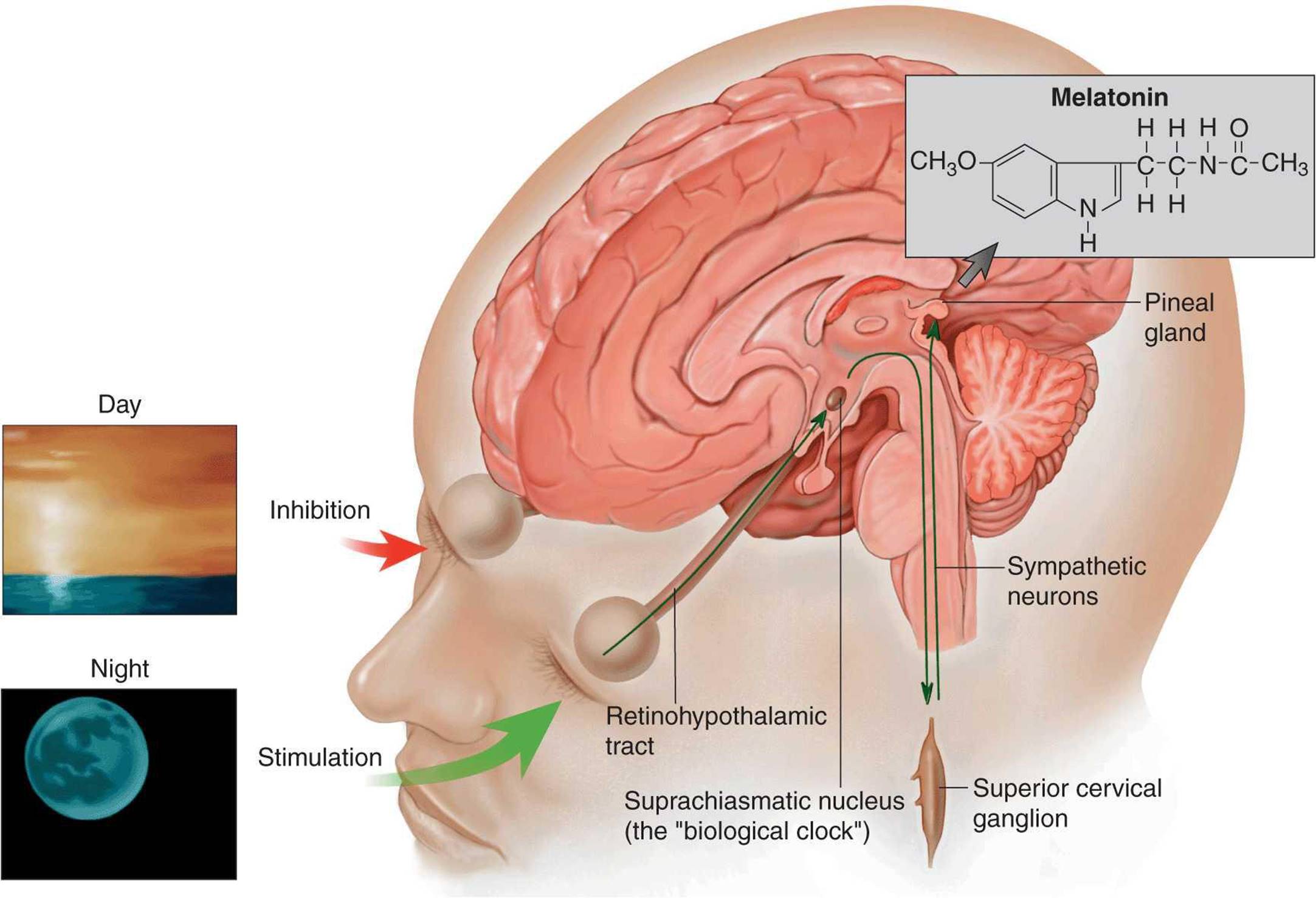
FIGURE 14–10 Secretion of melatonin. Retinohypothalamic fibers synapse in the suprachiasmatic nuclei (SCN), and there are connections from the SCN to sympathetic preganglionic neurons in the spinal cord that project to the superior cervical ganglion. Postganglionic neurons project from this ganglion to the pineal gland that secretes melatonin. The cyclic activity of SCN sets up a circadian rhythm for melatonin release. This rhythm is entrained to light/dark cycles by neurons in the retina. (From Fox SI: Human Physiology. McGraw-Hill, 2008.)
Evidence suggests that the SCN have two peaks of circadian activity. This may correlate with the observation that exposure to bright light can either advance, delay, or have no effect on the sleep–wake cycle in humans depending on the time of day when it is experienced. During the usual daytime it has no effect, but just after dark it delays the onset of the sleep period, and just before dawn it accelerates the onset of the next sleep period. Injections of melatonin have similar effects. In experimental animals, exposure to light turns on immediate-early genes in the SCN, but only at times during the circadian cycle when light is capable of influencing entrainment. Stimulation during the day is ineffective. Clinical Box 14–4 describes circadian rhythm disorders that impact the sleep–wake state.
CLINICAL BOX 14–4
Insomnia and Circadian Rhythm Disturbances of the Sleep–Wake State
Insomnia is defined as difficulty in initiating and/or maintaining sleep several times a week. Nearly 30% of adults report episodes of insomnia, and more than 50% of those aged 65 or older experience sleep problems. Individuals with persistent episodes of insomnia are more likely to experience accidents, a diminished work experience, and a poorer overall quality of life. Insomnia is often co-morbid with depression, and both disorders show abnormal regulation of corticotropin-releasing factor.
There are two major types of sleep disorders associated with disruption of the circadian rhythm. These are transient sleep disorders (jet lag, altered sleep cycle because of shift work, and illness) and chronic sleep disorders (delayed or advanced sleep phase syndrome). Those with delayed sleep phase syndrome have the inability to fall asleep in the evenings and awaken in the mornings. However, they have a normal total sleep time. Those with advanced sleep phase syndrome consistently fall asleep in early evening and awaken in early morning. This is seen primarily in the elderly and the depressed.
THERAPEUTIC HIGHLIGHTS
Light therapy has been shown to be effective to treat individuals who experience disturbances in their circadian cycle. Melatonin can be used to treat jet lag, and insomnia in elderly individuals. Ramelteon is a MT1 and MT2melatonin receptor agonist that is more effective than melatonin in treating insomnia. Zolpidem (ambien) is an example of a sedative-hypnotic that slows brain activity to promote sleep onset. In addition to treating daytime sleepiness in narcolepsy, modafinil has also been used successfully in the treatment of daytime sleepiness due to shift work and possibly to treat delayed sleep disorder syndrome.
NEUROCHEMICAL MECHANISMS PROMOTING SLEEP & AROUSAL
Transitions between sleep and wakefulness manifest a circadian rhythm consisting of an average of 6–8 h of sleep and 16–18 h of wakefulness. Nuclei in both the brain stem and hypothalamus are critical for the transitions between these states of consciousness. A classic study by Moruzzi and Magoun showed in 1949 that high-frequency stimulation of the midbrain reticular formation (the RAS) produces the EEG alerting response and arouses a sleeping animal. Damage to the area produces a comatose state. Electrical stimulation of the posterior hypothalamus also produces arousal similar to that elicited by stimulation of the midbrain, while electrical stimulation of the anterior hypothalamus and adjacent basal forebrain region induces sleep.
As described above, the brainstem RAS is composed of several groups of neurons that release norepinephrine, serotonin, or acetylcholine. The locations and wide projections of these neuronal populations are shown in Figure 7–2. In the case of the forebrain neurons involved in control of the sleep–wake cycles, preoptic neurons in the hypothalamus release GABA and posterior hypothalamic neurons release histamine. Also, orexin is produced in hypothalamic neurons and appear to be important in switching between sleep and wakefulness.
One theory regarding the basis for transitions from sleep to wakefulness involves alternating reciprocal activity of different groups of RAS neurons. In this model (Figure 14–11), wakefulness and REM sleep are at opposite extremes. When the activity of norepinephrine- and serotonin-containing neurons (locus coeruleus and raphé nuclei) is dominant, there is a reduced level of activity in acetylcholine-containing neurons in the pontine reticular formation. This pattern of activity contributes to the appearance of the awake state. The reverse of this pattern leads to REM sleep. When there is a more even balance in the activity of the aminergic and cholinergic neurons, NREM sleep occurs. The orexin released from hypothalamic neurons may regulate the changes in activity in these brainstem neurons.
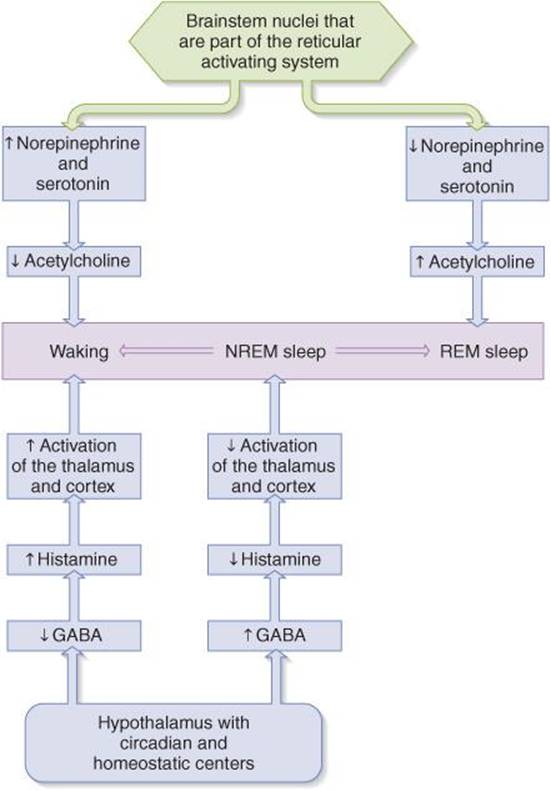
FIGURE 14–11 A model of how alternating activity of brain stem and hypothalamic neurons may influence the different states of consciousness. In this model, wakefulness and REM sleep are at opposite extremes. When the activity of norepinephrine- and serotonin-containing neurons (locus coeruleus and raphé nuclei) is dominant, there is a reduced level of activity in acetylcholine-containing neurons in the pontine reticular formation leading to wakefulness. The reverse of this pattern leads to REM sleep. A more even balance in the activity of these groups of neurons is associated with NREM sleep. Increases in GABA and decreases in histamine promote NREM sleep via deactivation of the thalamus and cortex. Wakefulness occurs when GABA is reduced and histamine is released. (From Widmaier EP, Raff H, Strang KT: Vander’s Human Physiology, 11th ed. McGraw-Hill, 2008.)
In addition, an increased release of GABA and reduced release of histamine increase the likelihood of NREM sleep via deactivation of the thalamus and cortex. Wakefulness occurs when GABA release is reduced and histamine release is increased.
MELATONIN AND THE SLEEP–WAKE STATE
In addition to the previously described neurochemical mechanisms promoting changes in the sleep–wake state, melatonin release from the richly vascularized pineal gland plays a role in sleep mechanisms (Figure 14–12). The pineal arises from the roof of the third ventricle in the diencephalon and is encapsulated by the meninges. The pineal stroma contains glial cells and pinealocytes with features suggesting that they have a secretory function. Like other endocrine glands, it has highly permeable fenestrated capillaries. In infants, the pineal is large and the cells tend to be arranged in alveoli. It begins to involute before puberty and small concretions of calcium phosphate and carbonate (pineal sand) appear in the tissue. Because the concretions are radiopaque, the pineal is often visible on x-ray films of the skull in adults. Displacement of a calcified pineal from its normal position indicates the presence of a space-occupying lesion such as a tumor in the brain.
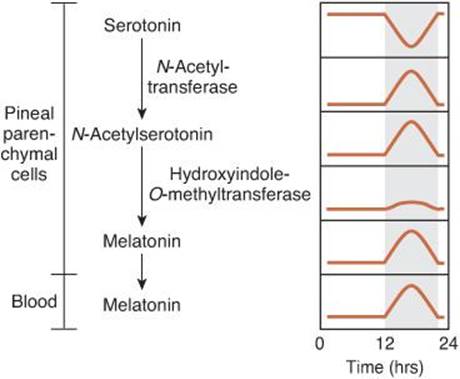
FIGURE 14–12 Diurnal rhythms of compounds involved in melatonin synthesis in the pineal. Melatonin and the enzymes responsible for its synthesis from serotonin are found in pineal pinealocytes; melatonin is secreted into the bloodstream. Melatonin synthesis and secretion are increased during the dark period (shaded area) and maintained at a low level during the light period.
Melatonin and the enzymes responsible for its synthesis from serotonin by N-acetylation and O-methylation are present in pineal pinealocytes, and the hormone is secreted by them into the blood and the cerebrospinal fluid (Figure 14–12). Two melatonin receptors (MT1 and MT2) have been found on neurons in the SCN. Both are G protein coupled receptors, with MT1 receptors inhibiting adenylyl cyclase and resulting in sleepiness. MT2 receptors stimulate phosphoinositide hydrolysis and may function in synchronization of the light—dark cycle.
The diurnal change in melatonin secretion may function as a timing signal to coordinate events with the light–dark cycle in the environment. Melatonin synthesis and secretion are increased during the dark period of the day and maintained at a low level during daylight hours (Figure 14–12). This diurnal variation in secretion is brought about by norepinephrine secreted by the postganglionic sympathetic nerves that innervate the pineal gland (Figure 14–10). Norepinephrine acts via β-adrenergic receptors to increase intracellular cAMP, and the cAMP in turn produces a marked increase in N-acetyltransferase activity. This results in increased melatonin synthesis and secretion. Circulating melatonin is rapidly metabolized in the liver by 6-hydroxylation followed by conjugation, and over 90% of the melatonin that appears in the urine is in the form of 6-hydroxy conjugates and 6-sulfatoxymelatonin. The pathway by which the brain metabolizes melatonin is unsettled but may involve cleavage of the indole nucleus.
The discharge of the sympathetic nerves to the pineal is entrained to the light–dark cycle in the environment via the retinohypothalamic nerve fibers to the SCN. From the hypothalamus, descending pathways converge onto preganglionic sympathetic neurons that in turn innervate the superior cervical ganglion, the site of origin of the postganglionic neurons to the pineal gland.
CHAPTER SUMMARY
![]() The major rhythms in the EEG are alpha (8–13 Hz), beta (13–30 Hz), theta (4–7 Hz), delta (0.5–4 Hz), and gamma (30–80 Hz) oscillations.
The major rhythms in the EEG are alpha (8–13 Hz), beta (13–30 Hz), theta (4–7 Hz), delta (0.5–4 Hz), and gamma (30–80 Hz) oscillations.
![]() The EEG is of some value in localizing pathologic processes, and it is useful in characterizing different types of seizures.
The EEG is of some value in localizing pathologic processes, and it is useful in characterizing different types of seizures.
![]() Throughout NREM sleep, there is some activity of skeletal muscle. A theta rhythm can be seen during stage 1 of sleep. Stage 2 is marked by the appearance of sleep spindles and occasional K complexes. In stage 3, a delta rhythm is dominant. Maximum slowing with slow waves is seen in stage 4. REM sleep is characterized by low-voltage, high-frequency EEG activity and rapid, roving movements of the eyes.
Throughout NREM sleep, there is some activity of skeletal muscle. A theta rhythm can be seen during stage 1 of sleep. Stage 2 is marked by the appearance of sleep spindles and occasional K complexes. In stage 3, a delta rhythm is dominant. Maximum slowing with slow waves is seen in stage 4. REM sleep is characterized by low-voltage, high-frequency EEG activity and rapid, roving movements of the eyes.
![]() A young adult typically passes through stages 1 and 2, and spends 70–100 min in stages 3 and 4. Sleep then lightens, and a REM period follows. This cycle repeats at 90-min intervals throughout the night. REM sleep occupies 50% of total sleep time in full-term neonates; this proportion declines rapidly and plateaus at about 25% until it falls further in old age.
A young adult typically passes through stages 1 and 2, and spends 70–100 min in stages 3 and 4. Sleep then lightens, and a REM period follows. This cycle repeats at 90-min intervals throughout the night. REM sleep occupies 50% of total sleep time in full-term neonates; this proportion declines rapidly and plateaus at about 25% until it falls further in old age.
![]() Transitions from sleep to wakefulness may involve alternating reciprocal activity of different groups of RAS neurons. When the activity of norepinephrine- and serotonin-containing neurons is dominant, the activity in acetylcholine-containing neurons is reduced, leading to the appearance of wakefulness. The reverse of this pattern leads to REM sleep. Also, wakefulness occurs when GABA release is reduced and histamine release is increased.
Transitions from sleep to wakefulness may involve alternating reciprocal activity of different groups of RAS neurons. When the activity of norepinephrine- and serotonin-containing neurons is dominant, the activity in acetylcholine-containing neurons is reduced, leading to the appearance of wakefulness. The reverse of this pattern leads to REM sleep. Also, wakefulness occurs when GABA release is reduced and histamine release is increased.
![]() The entrainment of biological processes to the light–dark cycle is regulated by the SCN.
The entrainment of biological processes to the light–dark cycle is regulated by the SCN.
![]() The diurnal change in melatonin secretion from serotonin in the pineal gland may function as a timing signal to coordinate events with the light–dark cycle, including the sleep–wake cycle.
The diurnal change in melatonin secretion from serotonin in the pineal gland may function as a timing signal to coordinate events with the light–dark cycle, including the sleep–wake cycle.
MULTIPLE-CHOICE QUESTIONS
For all questions, select the single best answer unless otherwise directed.
1. In a healthy, alert adult sitting with their eyes closed, the dominant EEG rhythm observed with electrodes over the occipital lobes is
A. delta (0.5–4 Hz).
B. theta (4–7 Hz).
C. alpha (8–13 Hz).
D. beta (18–30 Hz).
E. fast, irregular low-voltage activity.
2. A 35-year-old male spent the evening in a sleep clinic to determine whether he had obstructive sleep apnea. The tests showed that NREM sleep accounted for over 30% of his total sleep time. Which of the following pattern of changes in central neurotransmitters or neuromodulators are associated with the transition from NREM to wakefulness?
A. Decrease in norepinephrine, increase in serotonin, increase in acetylcholine, decrease in histamine, and decrease in GABA.
B. Decrease in norepinephrine, increase in serotonin, increase in acetylcholine, decrease in histamine, and increase in GABA.
C. Decrease in norepinephrine, decrease in serotonin, increase in acetylcholine, increase in histamine, and increase in GABA.
D. Increase in norepinephrine, increase in serotonin, decrease in acetylcholine, increase in histamine, and decrease in GABA.
E. Increase in norepinephrine, decrease in serotonin, decrease in acetylcholine, increase in histamine, and decrease in GABA.
3. A gamma rhythm (30–80 Hz)
A. is characteristic of seizure activity.
B. is seen in an individual who is awake but not focused.
C. may be a mechanism to bind together sensory information into a single percept and action.
D. is independent of thalamocortical loops.
E. is generated in the hippocampus.
4. For the past several months, a 67-year-old female experienced difficulty initiating and/or maintaining sleep several times a week. A friend suggested that she take melatonin to regulate her sleep–wake cycle. Melatonin secretion would probably not be increased by
A. stimulation of the superior cervical ganglia.
B. intravenous infusion of tryptophan.
C. intravenous infusion of epinephrine.
D. stimulation of the optic nerve.
E. induction of pineal hydroxyindole-O-methyltransferase.
5. A 10-year-old boy was diagnosed with childhood absence epilepsy. His EEG showed a bilateral synchronous, symmetrical 3-Hz spike-and-wave discharge. Absence seizures
A. are a form of nonconvulsive generalized seizures accompanied by momentary loss of consciousness.
B. are a form of complex partial seizures accompanied by momentary loss of consciousness.
C. are a form of nonconvulsive generalized seizures without a loss of consciousness.
D. are a form of simple partial seizures without a loss of consciousness.
E. are a form of convulsive generalized seizures accompanied by momentary loss of consciousness.
6. A 57-year-old professor at a medical school experienced numerous episodes of a sudden loss of muscle tone and an irresistible urge to sleep in the middle of the afternoon. He was diagnosed with narcolepsy which
A. is characterized by a sudden onset of NREM sleep.
B. has a familial incidence associated with a class II antigen of the major histocompatibility complex.
C. may be due to the presence of an excessive number of orexin-producing neurons in the hypothalamus.
D. is often effectively treated with dopamine receptor agonists.
E. is the most common cause of daytime sleepiness.
CHAPTER RESOURCES
Blackman S: Consciousness: An Introduction. Oxford University Press, 2004.
Feely M: Drug treatment of epilepsy. British Med J 1999;318:106.
McCormick DA, Contreras D: Of the cellular and network bases of epileptic seizures. Annu Rev Physiol 2001;63:815.
Merica H, Fortune RD: State transitions between wake and sleep, and within the ultradian cycle, with focus on the link to neuronal activity. Sleep Med Rev 2004;8:473.
Oberheim NA, Tian GF, Han X, et al: Loss of astrocytic domain organization in the epileptic brain. J Neurosci 2008;28:3264.
Sakurai T: The neural circuit of orexin (hypocretin): maintaining sleep and wakefulness. Nature Rev Neurosci 2007;8:171.
Saper CB, Fuller PM, Pedersen NP, Lu J, Scrammell TE: Sleep state switching. Neuron 2010;68:1023.
Shaw JC (editor): The Brain’s Alpha Rhythms and the Mind. Elsevier, 2003.
Siegel JM: Narcolepsy. Sci Am 2000;282:76.
Stafstrom CE: Epilepsy: A review of selected clinical syndromes and advances in basic science. J Cereb Blood Flow Metab 2006;26:983.
Steinlein O: Genetic mechanisms that underlie epilepsy. Nat Rev Neurosci 2004;5:400.
Steriade M, McCarley RW: Brain Stem Control of Wakefulness and Sleep. Plenum, 1990.
Steriade M, Paré D: Gating in Cerebral Networks. Cambridge University Press, 2007.
Thorpy M (editor): Handbook of Sleep Disorders. Marcel Dekker, 1990.