Anne Schumacher1 and Ana Claudia Zenclussen1
(1)
Department of Experimental Obstetrics and Gynecology, Medical Faculty, Otto-von-Guericke University, Gerhart-Hauptmann Straße 35, 39108 Magdeburg, Germany
Ana Claudia Zenclussen
Email: ana.zenclussen@med.ovgu.de
Abstract
Recognition of foreign paternal antigens expressed in the semi-allogeneic fetus by maternal immune cells is a requirement for successful pregnancy. However, despite intensive research activity during the last decades, the precise mechanisms contributing to the acceptance of the paternal alloantigens are still puzzling and pregnancy remains a fascinating phenomenon. Moreover, most studies focused on the maternal and fetal contribution to pregnancy success, and relatively little is known about the paternal involvement. In the current review, we address the contribution of paternal-derived factors to fetal-tolerance induction. First, we discuss data suggesting that in both humans and mice, the female body gets prepared for a pregnancy in every cycle, also in regard to male alloantigens delivered at coitus. Then, we provide an overview about factors present in seminal fluid and how these factors influence immune responses in the female reproductive tract. We further discuss ways of paternal alloantigen presentation and identify the immune modulatory properties of seminal fluid-derived factors with a special focus on Treg biology. Finally, we highlight the therapeutic potential of seminal fluid in different clinical applications.
Keywords
AlloantigensFetal toleranceRegulatory T cellsSeminal fluidSeminal plasmaPregnancy
10.1 Introduction
Awareness of the foreign paternal/fetal alloantigens and their tolerance is a prerequisite for the survival of the semi-allogeneic fetus within the maternal uterus. There is plenty of published evidence that, to achieve this tolerance, both mother and fetus release immune-modulating factors that among other effects augment the number of regulatory immune cells to suppress alloreactive immune responses. By contrast, the contribution of the father to fetal tolerance is still underexplored. When a couple plans to have a baby, they should have sexual intercourse without contraception. Within this time, even if pregnancy does not occur, maternal immune cells present in the vagina and the uterus are repeatedly challenged by paternal alloantigens present in the semen at coitus. Several studies indicate that there is an accumulation of various immune cell populations in the vagina and the uterus at or immediately after ovulation, and these cellular changes are needed to establish fetal tolerance from the very beginning of pregnancy. Early recognition and acceptance of paternal alloantigens has been shown to ensure successful embryo implantation and fetal growth. Preeclampsia (PE) is a serious pregnancy complication associated with poor placentation as a consequence of shallow trophoblast invasion. The risk of developing PE is diminished if repeated contact with paternal alloantigens has taken place before pregnancy arises (Saftlas et al. 2014). Dekker and colleagues discuss in their review that PE is a disease of first pregnancies. The protective effect of multiparity is however lost with change of partner. Artificial donor insemination and oocyte donation result in a substantial increase of PE risk (Dekker et al. 1998). These observations underlie the meaningfulness of early alloantigen awareness for a successful progress of pregnancy as immune maladaptation is related with poor pregnancy outcome. We need to further study mechanisms underlying establishment of early fetal allotolerance. Here, we discuss available data from the literature on the immune-modulating properties of seminal fluid components with a specific focus on regulatory T cells (Treg).
10.2 Immune Cell Fluctuations During the Reproductive Cycle
Fluctuations in various immune cell populations have been observed in uterine tissue during the reproductive cycle in humans and mice. These cellular changes are proposed to prepare the endometrium for the appearance of fetal alloantigens when the embryo is implanting. Hormonal variations and other factors have been held responsible for the observed fluctuations. For instance, uterine natural killer (NK) cells dramatically increase in number in the late secretory phase of the human menstrual cycle finally representing the main immune cell population of all leukocytes in human decidual tissue (Bulmer et al. 1991). Moreover, human and murine uterine mast cells (MCs) oscillate during the reproductive cycle (Padilla et al. 1990; Mori et al. 1997) reaching their maximum number in the receptive phase (estrus) of the murine estrous cycle (Woidacki et al. 2013). In line, we demonstrated that dendritic cells (DCs) and Treg accumulate in the estrus phase in mice (Zenclussen et al. 2013; Teles et al. 2013a). For the latter, cycle-dependent fluctuations have been confirmed also by others in human and mice (Arruvito et al. 2007; Kallikourdis and Betz 2007), and it was proposed that Treg changes are hormone driven (Weinberg et al. 2011; Schumacher et al. 2014). Altogether, it can be assumed that every time a female becomes receptive, her immune system prepares itself for the contact with the foreign paternal antigens delivered with semen at coitus. This ensures the possibility of a very early tolerance initiation toward the paternal antigens that will be presented by fetal tissue at implantation and afterward.
10.3 Composition of Seminal Fluid
Seminal fluid (semen) consists of spermatozoa suspended in seminal plasma. Components of seminal plasma are secreted from rete testis, epididymis, and accessory sex glands including the seminal vesicle, the prostate, and the bulbourethral glands (Juyena and Stelletta 2012). Analysis of mammalian seminal plasma revealed a variety of factors including ions, energy substrates, organic compounds, and nitrogenous compounds (Juyena and Stelletta 2012; Milardi et al. 2013). Additionally, seminal plasma contains high concentrations of factors proven to posses immune regulatory properties. One of these factors is tumor growth factor-β (TGF-β), whereby TGF-β1 and TGF-β3 can be found in high amounts and TGF-β2 is present in lower amounts (Nocera and Chu 1995; Lokeshwar and Block 1992; O’Leary et al. 2013; Srivastava et al. 1996; Tremellen et al. 1998). Therefore, the majority of TGF-β existing in its latent form has to be activated to be fully functional. TGF-β activation was proposed to take place after insemination in the female tract. Here, several enzymes delivered with the seminal plasma or being present in the female tract are suggested to contribute to the activation process. More precisely, plasmin, subtilisin-like endoproteases, tissue- and urokinase-type plasminogen activator (Chu and Kawinski 1998), thrombospondin 1 (Slater and Murphy 1999), and αvβ6 integrin (Breuss et al. 1993) were implicated in TGF-β activation. Moreover, the acidic environment of the human vagina may drive TGF-β activation to some extent. Besides TGF-β, human seminal plasma contains other immune-modulating factors such as interleukin (IL)-8 and soluble IL-2 receptor (Srivastava et al. 1996) prostaglandin E2 (PGE2) and 19-hydroxyprostaglandin E (19-hydroxy PGE) (Denison et al. 1999), soluble tumor necrosis factor (TNF) receptors (Liabakk et al. 1993), receptors for the Fc portion of γ-globulin, spermine (Evans et al. 1995), and complement inhibitors (Kelly 1995).
10.4 Recognition and Presentation of Seminal Fluid-Derived Alloantigens
Immediately after insemination, the secretion of growth factors, cytokines, and chemokines from cervical and endometrial tissue is induced, resulting in local pro-inflammatory environment. This provokes a rapid and dramatic influx of immune cells and additionally increases the number of various immune cell populations in situ. In the mouse, release of the granulocyte-macrophage colony-stimulating factor (GM-CSF), IL-6, and several chemokines leads to the infiltration of macrophages, DCs, and granulocytes in subepithelial stromal tissue (De et al. 1991; McMaster et al. 1992; Robertson et al. 1996). In humans, sexual intercourse results in neutrophil infiltration into the superficial epithelium of the cervical tissues (Pandya and Cohen 1985) followed by a recruitment of macrophages, DCs, and lymphocytes into the epithelial layers and deeper stromal tissues (Sharkey et al. 2007). This remarkable recruitment of immune cells and especially of those capable of efficiently present alloantigens suggests that presentation of paternal antigens can take place even before implantation and be involved in pregnancy success or failure already at this early time point. Evidence for this was provided by two studies showing that depletion of Treg before mating or depletion of uterine DCs in the preimplantation period impaired the implantation process drastically (Plaks et al. 2008; Teles et al. 2013a). We confirmed the presence of paternal alloantigens immediately after fecundation in a mouse model. By mating wild-type females with GFP+ males, we demonstrated that paternal antigens (GFP+ cells) can be found in vaginal lumen as early as day 0.5 of pregnancy. Moreover, at the same time, we detected GFP+ cells in decidual tissue and lymph nodes. At later pregnancy stages, we found GFP+ cells in lymphoid as well as nonlymphoid tissue. However, after day 5 of murine pregnancy (implantation), we could not distinguish whether GFP+ structures were of paternal or fetal origin (Zenclussen et al. 2010). The appearance of fetal cells in maternal organs as a result of transplacental cell migration is well documented in humans and mice and results in a phenomenon called fetal microchimerism (Yan et al. 2005; Khosrotehrani et al. 2005; Tan et al. 2005). Accordingly, maternal cells migrate to the fetus ending up in maternal microchimerism (Loubière et al. 2006). In more detailed analysis, we then showed that some GFP+ cells also expressed MHC class II molecules and were positive for the DC marker CD11c suggesting that semen contains DCs capable to present paternal antigens immediately after fecundation. In line with these findings, Witkin and colleagues confirmed the presence of several immune cell populations in semen (Witkin and Goldstein 1988). Together with the observation that maternal antigen-presenting cells (APCs) are enriched in the vagina and uterus after insemination, it can be assumed that paternal alloantigens are presented directly (via paternal APCs) and/or indirectly (via maternal APCs). Moldenhauer and colleagues analyzed the involvement of seminal fluid antigens in maternal T-cell activation and defined the underlying antigen- presenting pathway in a transgenic mouse model. The authors transferred T cells specific for the ovalbumin antigen to female mice that were mated to male mice ubiquitously expressing membrane-bound ovalbumin. They confirmed the presence of the ovalbumin antigen in the seminal plasma and detected activated ovalbumin-specific CD4+ and CD8+ T cells locally in the para-aortic lymph nodes displaying a high proliferative capacity. Furthermore, they proved that seminal plasma, but not sperm, was necessary to induce T-cell proliferation. Interestingly, they showed that paternal ovalbumin presented by maternal APCs was essential for CD8+ T-cell proliferation, but responses were not elicited when ovalbumin was presented by paternal APCs (Moldenhauer et al. 2009) suggesting that indirect rather that direct pathways are involved. However, two other groups proposed antigen presentation via the direct pathway (Root-Bernstein and DeWitt 1995; Clark et al. 2013). It was suggested that the direct presentation of paternal antigens may also determine the occurrence of spontaneous abortion events (Clark et al. 2013). More studies are necessary to clarify the pathways of paternal antigens presentation and their relevance for pregnancy success.
10.5 Immune Regulatory Properties of Seminal Plasma
Sarah Robertson and colleagues discussed four different effector functions mediated by seminal plasma on reproductive processes (Robertson 2005). These functions include the clearance of sperms and microbes delivered with semen at mating, the activation of tissue remodeling processes essential for embryo implantation, the induction of cytokines and growth factors important for preimplantation embryo development, and the induction of tolerance mechanisms toward the foreign paternal alloantigens. Additionally, very recently, the same research group provided important evidence that seminal plasma can also affect the health of male offsprings. By ablation of the plasma fraction from seminal fluid, the authors observed male offsprings exhibiting obesity, distorted metabolic hormones, diminished glucose tolerance, and hypertension suggesting that seminal plasma has long-lasting effects on the health of the male progenies (Bromfield et al. 2014).
Here, we will focus on the immune-modulating properties of seminal plasma components. As one of the major components, TGF-β is able to negatively influence growth activity and function of lymphocytes (Nocera and Chu 1993). Moreover, together with IL-8, TGF-β has been shown to induce IL-1β, IL-6, and LIF expression in endometrial epithelial cells (Gutsche et al. 2003). LIF itself was reported to play an essential role for embryo implantation and development, and its lack has been associated with pregnancy loss in the mouse (Stewart et al. 1992). Whether this is relevant for human pregnancies, it is still a matter of debate. Furthermore, other factors present in seminal plasma were reported to influence immune responses, for instance, the soluble p55 TNF-α receptor known to inhibit TNF-mediated cytotoxicity (Liabakk et al. 1993), spermine which impairs proliferation of NK cells and T lymphocytes (Evans et al. 1995), and inhibitors of the complement system (Tarter and Alexander 1984; Chowdhury et al. 1996). Finally, prostaglandins were suggested to prevent lymphocyte proliferation, NK cell activity, and secretion of pro-inflammatory cytokines (Kelly 1995; Kelly et al. 1997).
10.6 Regulatory T Cells in Human and Murine Pregnancy
Treg represent a unique T-cell subpopulation best known for their function in suppressing autoreactive and alloreactive immune responses, thereby preventing autoimmune diseases and allograft rejection (Sakaguchi et al. 1995). In addition, a crucial role for Treg in the establishment and maintenance of fetal tolerance has widely been reported in both humans and mice (Aluvihare et al. 2004; Zenclussen et al. 2005; Heikkinen et al. 2004; Saito et al. 2005). Most studies observed an augmentation of Treg in peripheral blood and decidua during the first and second trimester in normal pregnant women (Heikkinen et al. 2004; Xiong et al. 2013; Tilburgs et al. 2006). In the third trimester, Treg levels begin to decrease (Seol et al. 2008) and further decline with successive stages of labor (Xiong et al. 2010). Furthermore, Treg suppressive capacity is reduced in term labor and preterm labor suggesting that changes in Treg function may contribute to initiation of labor (Kisielewicz et al. 2010). In mice, normal pregnancy is associated with an augmentation in Treg numbers at very early pregnancy stages, a reduction around implantation time followed by a second increment on day 10 of pregnancy (Teles et al. 2013b; Thuere et al. 2007). Uterine enrichment of Treg can be achieved by a selective recruitment of these cells from the periphery or by conversion from conventional T cells directly at the fetal-maternal interface (Ramhorst et al. 2012; Tilburgs et al. 2008). Cytokines, chemokines, and hormones may serve as attractors for Treg migration into uterine tissue (Kallikourdis et al. 2007; Schumacher et al. 2009; Teles et al. 2013a). We recently suggested that CCR7 is involved in murine Treg homing to nonpregnant uterus (Teles et al. 2013a), while the pregnancy-hormone human chorionic gonadotropin attracts human Treg to trophoblasts (Schumacher et al. 2009) and murine Treg to the fetal-maternal interface when injected at peri-implantation (Schumacher et al. 2013). LH also attracted Treg to the fetal-maternal interface in a mouse model (Schumacher et al. 2014).
The importance of Treg for pregnancy success was confirmed by several studies showing that in both humans and mice, spontaneous abortion is associated with a diminished number and activity of Treg (Aluvihare et al. 2004; Zenclussen et al. 2005; Heikkinen et al. 2004; Somerset et al. 2004; Sasaki et al. 2004; Yang et al. 2008; Jin et al. 2009; Mei et al. 2010; Inada et al. 2013). Additionally, reduced Treg levels and impaired functionality were reported in other pregnancy complications such as extrauterine pregnancies, endometriosis, and preeclampsia (Schumacher et al. 2009; Basta et al. 2010; Sasaki et al. 2007; Toldi et al. 2008; Prins et al. 2009; Santner-Nanan et al. 2009; Quinn et al. 2011; Darmochwal-Kolarz et al. 2012), clearly indicating that only fully functional Treg guarantee successful pregnancy. The adoptive transfer of Treg was shown to diminish the occurrence of spontaneous abortion in the mouse (Zenclussen et al. 2005; Schumacher et al. 2007; Yin et al. 2012). Immunization with paternal antigens in early human pregnancies was associated with an increase of Treg (Wu et al. 2014).
10.7 Antigen Specificity of Treg and Mechanisms of Their Action
There is a general consent that recognition of fetal alloantigens by maternal immune cells is a prerequisite for the induction of an active suppression of anti-fetal immune responses. However, there is still a lively debate whether Treg mediate their protection in an antigen-specific fashion. Although several studies indicated that the recognition of foreign paternal/fetal alloantigens is critically for Treg development and function (Darrasse-Jèze et al. 2006; Mjösberg et al. 2007; Kallikourdis et al. 2007; Tilburgs et al. 2009; Kahn and Baltimore 2010; Schumacher et al. 2007; Liu et al. 2013), there is also evidence arguing against a “pure” alloantigen-driven expansion of Treg during pregnancy (Chen et al. 2013). Based on these findings, it seems presumable that both alloantigens and self-antigens support Treg augmentation and protective activity during pregnancy.
In contrast to the discussed antigen specificity of Treg function, the research community agrees on the indispensable role for Treg in the prevention of overwhelming inflammatory immune responses in various clinical disciplines. Here, Treg have been reported to modulate the number and activity of every other immune cell population. For instance, Treg suppress the proliferation and function of T cells, DCs, and macrophages (Piccirillo and Shevach 2001; Mempel et al. 2006; Cederbom et al. 2000; Misra et al. 2004; Taams et al. 2005) and hamper the proliferation of B cells as well as their antibody secretion (Lim et al. 2005). Treg function is thereby realized either by direct cell-cell contact or via the secretion of immune-suppressive cytokines such as IL-10 and TGF-β (Hara et al. 2001; Wahl et al. 2004; Friedline et al. 2009). In pregnancy, the precise mechanisms underlying Treg protection are still not completely understood. We found that in a murine model of disturbed fetal tolerance, Treg rather function through PD-1, IL-10, and HO-1 than through CTLA-4 and TGF-β (Verdijk et al. 2004; Schumacher et al. 2007, 2012). However, Jin and colleagues proposed a role for CTLA-1 in Treg function in humans (Jin et al. 2009), suggesting that the way of Treg-mediated suppression may differ between experimental models and species. Additionally, we recently showed that Treg regulate the accumulation of conventional CD8+T cells and the production of pro-inflammatory molecules in the uterus and draining lymph nodes in the preimplantation period. Therefore, Treg dampen local inflammatory processes occurring during the time of implantation and support successful embryo nidation (Teles et al. 2013a).
10.8 Influence of Seminal Fluid on Treg Biology
There is accumulating evidence that seminal fluid-derived antigens and other factors present in seminal fluid play a pivotal role in Treg generation, expansion, migration, and function. In the preimplantation period, seminal fluid causes an expansion of the CD4+CD25+Foxp3+ Treg population in uterine-draining lymph nodes and the uterus itself. The need for seminal fluid for uterine Treg expansion was proven by matings with seminal vesicle-deficient and vasectomized males (Guerin et al. 2011; Robertson et al. 2009) suggesting that both seminal plasma and sperms are necessary for Treg induction. We confirmed the need for seminal plasma for in vivo Treg expansion in draining lymph nodes in the preimplantation period by matings with seminal-deficient male mice (Teles et al. 2013a). In addition, in vitro co-culture of Treg in the presence of different concentrations of seminal plasma provoked a significant proliferation of Treg. However, this was not observed for conventional T cells (Teles et al. 2013a). Moreover, pseudopregnancy induced by mechanical stimulation did not result in Treg augmentation again underlying the importance of seminal plasma for Treg elevation (Schumacher et al. 2007). In contrast to observations obtained by Robertson and colleagues, in our mouse model, matings with vasectomized males did not significantly impair Treg increase in lymph nodes (Schumacher et al. 2007; Teles et al. 2013a).
TGF-β, present in high amounts in seminal plasma, might be one of the factors responsible for seminal plasma-driven Treg expansion. In vitro co-cultures of seminal plasma with Treg in the presence of a TGF-β antibody abrogated Treg proliferation (Teles et al. 2013a). In line, Clark and colleagues confirmed an effect of pure TGF-β3 on Treg induction and pregnancy success. They applied pharmaceutical-grade bioactive TGF-β3 into the vaginal tract of abortion-prone females at mating. Application of intravaginal TGF-β3 reduced the abortion rate and increased the numbers of Treg in the vagina (Clark et al. 2008). PGE, known to have the capability to induce Treg number and activity (Baratelli et al. 2005), may also contribute to Treg elevation. Furthermore, seminal fluid induces the expression of uterine CCL19, a chemokine that acts through the CCR7 receptor, and may therefore be involved in Treg recruitment to the uterus (Guerin et al. 2011; Teles et al. 2013a). Moreover, Robertson and colleagues proved that seminal fluid-induced Treg are fully functional. In a mouse model, they nicely showed that Treg induced by seminal fluid-derived paternal antigens efficiently prevented the rejection of engrafted tumor cells expressing the same paternal antigens (Robertson et al. 2009). In agreement with the murine data, human seminal plasma was also shown to increase the proportion of CD127low CD49dlow Treg. However, Balandya and colleagues revealed that increased Treg numbers were a result of an increased conversion rate from CD4+ non-Treg into Foxp3- Treg and not due to proliferation of preexisting Treg (Balandya et al. 2012).
Altogether, Treg expansion in the preimplantation period may occur due to the presence of paternal alloantigens and other factors in seminal fluid. Paternal alloantigens might be presented by APCs of paternal or maternal origin. After implantation, the Treg pool is then further maintained by the continuous release of fetal antigens from the placenta supporting fetal survival until birth. Interestingly, postpartum fetal-specific Treg may persist in the mother creating a memory to paternal antigens (Schober et al. 2012) and rapidly re-accumulate during subsequent pregnancies (Rowe et al. 2012).
10.9 Therapeutic Potential of Seminal Fluid
The findings discussed above provide evidence that seminal fluid might possess some therapeutic potential in the treatment of infertility and miscarriage. A recently published review compared data on the outcome of in vitro fertilization (IVF) treatments in patients exposed to seminal plasma around the time of oocyte retrieval or embryo transfer with placebo controls or controls with no exposure to seminal plasma. The authors compared the clinical pregnancy and live birth/ongoing pregnancy rate and found a statistically significant improvement in clinical pregnancy rate but no significant improvement in terms of ongoing pregnancy/live birth rates. However, they admitted that available data for the ongoing pregnancy/live birth rates were very limited, and the methodology and quality of the analyzed studies were variable (Crawford et al. 2014). The positive effect of seminal plasma exposure on the clinical pregnancy rate in IVF patients might be explained by an increase in the number of Treg that has been associated with improved pregnancy rates in IVF patients (Zhou et al. 2012). Moreover, seminal fluid-driven Treg protection may also have beneficial effects in other clinical applications. In this regard, exposure of seminal fluid may improve disease activity in autoimmune disorders associated with reduced Treg numbers and function (Pakravan et al. 2014). Altogether, it can be assumed that administration of seminal fluid is a promising tool to modulate undesired immune responses and provoke tolerance in different clinical disciplines.
10.10 Conclusions
Despite intensive research work investigating the factors and mechanisms allowing fetal survival within the hostile uterine environment, there remain several open questions to be answered. One of these questions addresses the father’s contribution to fetal tolerance. Here, we pointed out that the father not only provides the genetics to create new life but also contributes to its survival from the very beginning by delivering immune-modulating factors with the semen. These factors allow the establishment and maintenance of a fetal-friendly environment resulting in a successful embryo implantation and fetal growth. Moreover, the health of the progeny after birth seems also to be influenced by paternal-derived factors. Thus, the contribution of the father for the survival of its own child should not be underestimated. In Fig. 10.1, we propose an hypothetical scenario as to how maternal immune responses are regulated upon contact with paternal antigens.
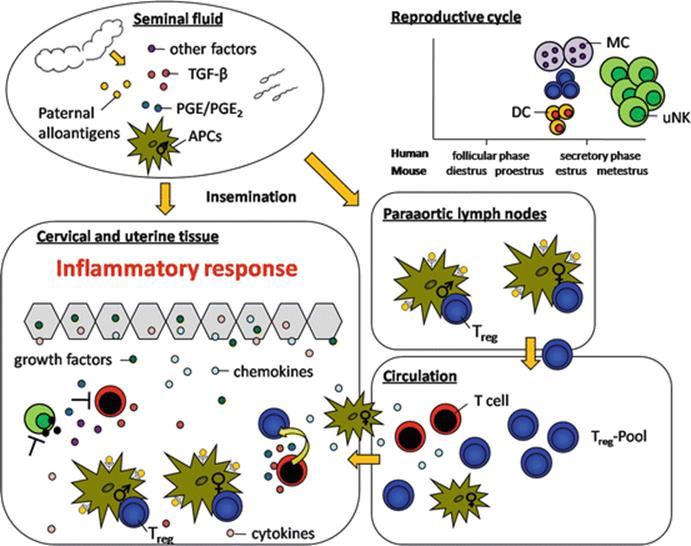
Fig. 10.1
Hypothetical scenario on the immune modulatory capacity of seminal fluid. After contact with paternal antigens a rather tolerant immune response is generated. The graphic illustrates the interactions between immune cells at different compartments
References
Aluvihare VR, Kallikourdis M, Betz AG (2004) Regulatory T cells mediate maternal tolerance to the fetus. Nat Immunol 5(3):266–271. doi:10.1038/ni1037PubMed
Arruvito L, Sanz M, Banham AH, Fainboim L (2007) Expansion of CD4+CD25+ and FOXP3+ regulatory T cells during the follicular phase of the menstrual cycle: implications for human reproduction. J Immunol 178(4):2572–2578PubMed
Balandya E, Wieland-Alter W, Sanders K, Lahey T (2012) Human seminal plasma fosters CD4+ regulatory T-cell phenotype and transforming growth factor-β1 expression. Am J Reprod Immunol 68(4):322–330. doi:10.1111/j.1600-0897.2012.01176.xPubMedCentralPubMed
Baratelli F, Lin Y, Zhu L, Yang S, Heuzé-Vourc'h N, Zeng G, Reckamp K, Dohadwala M, Sharma S, Dubinett SM (2005) Prostaglandin E2 induces FOXP3 gene expression and T regulatory cell function in human CD4+ T cells. J Immunol 175(3):1483–1490PubMed
Basta P, Majka M, Jozwicki W, Lukaszewska E, Knafel A, Grabiec M, Stasienko E, Wicherek L (2010) The frequency of CD25+CD4+ and FOXP3+ regulatory T cells in ectopic endometrium and ectopic decidua. Reprod Biol Endocrinol 8:116. doi:10.1186/1477-7827-8-116PubMedCentralPubMed
Breuss JM, Gillett N, Lu L, Sheppard D, Pytela R (1993) Restricted distribution of integrin beta 6 mRNA in primate epithelial tissues. J Histochem Cytochem 41(10):1521–1527PubMed
Bromfield JJ, Schjenken JE, Chin PY, Care AS, Jasper MJ, Robertson SA (2014) Maternal tract factors contribute to paternal seminal fluid impact on metabolic phenotype in offspring. Proc Natl Acad Sci 111(6):2200–2205. doi:10.1073/pnas.1305609111PubMedCentralPubMed
Bulmer JN, Morrison L, Longfellow M, Ritson A, Pace D (1991) Granulated lymphocytes in human endometrium: histochemical and immunohistochemical studies. Hum Reprod 6(6):791–798PubMed
Cederbom L, Hall H, Ivars F (2000) CD4+CD25+ regulatory T cells down-regulate co-stimulatory molecules on antigen-presenting cells. Eur J Immunol 30(6):1538–1543. doi:10.1002/1521-4141(200006)30:6<1538:AID-IMMU1538>3.0.CO;2-XPubMed
Chen T, Darrasse-Jeze G, Bergot A, Courau T, Churlaud G, Valdivia K, Strominger JL, Ruocco MG, Chaouat G, Klatzmann D (2013) Self-specific memory regulatory T cells protect embryos at implantation in mice. J Immunol 191(5):2273–2281. doi:10.4049/jimmunol.1202413PubMedCentralPubMed
Chowdhury NA, Kamada M, Takikawa M, Mori H, Gima H, Aono T (1996) Complement-inhibiting activity of human seminal plasma and semen quality. Arch Androl 36(2):109–118PubMed
Chu TM, Kawinski E (1998) Plasmin, subtilisin-like endoproteases, tissue plasminogen activator, and urokinase plasminogen activator are involved in activation of latent TGF-beta 1 in human seminal plasma. Biochem Biophys Res Commun 253(1):128–134. doi:10.1006/bbrc.1998.9760PubMed
Clark DA, Fernandes J, Fernandez J, Banwatt D (2008) Prevention of spontaneous abortion in the CBA x DBA/2 mouse model by intravaginal TGF-beta and local recruitment of CD4+8+ FOXP3+ cells. Am J Reprod Immunol 59(6):525–534. doi:10.1111/j.1600-0897.2008.00582.xPubMed
Clark DA, Rahmati M, Gohner C, Bensussan A, Markert UR, Chaouat G (2013) Seminal plasma peptides may determine maternal immune response that alters success or failure of pregnancy in the abortion-prone CBAxDBA/2 model. J Reprod Immunol 99(1–2):46–53. doi:10.1016/j.jri.2013.03.006PubMed
Crawford G, Ray A, Gudi A, Shah A, Homburg R (2014) The role of seminal plasma for improved outcomes during in vitro fertilization treatment: review of the literature and meta-analysis. Hum Reprod Update. doi:10.1093/humupd/dmu052PubMed
Darmochwal-Kolarz D, Kludka-Sternik M, Tabarkiewicz J, Kolarz B, Rolinski J, Leszczynska-Gorzelak B, Oleszczuk J (2012) The predominance of Th17 lymphocytes and decreased number and function of Treg cells in preeclampsia. J Reprod Immunol 93(2):75–81. doi:10.1016/j.jri.2012.01.006PubMed
Darrasse-Jèze G, Darasse-Jèze G, Klatzmann D, Charlotte F, Salomon BL, Cohen JL (2006) CD4+CD25+ regulatory/suppressor T cells prevent allogeneic fetus rejection in mice. Immunol Lett 102(1):106–109. doi:10.1016/j.imlet.2005.07.002PubMed
De M, Choudhuri R, Wood GW (1991) Determination of the number and distribution of macrophages, lymphocytes, and granulocytes in the mouse uterus from mating through implantation. J Leukoc Biol 50(3):252–262PubMed
Dekker GA, Robillard PY, Hulsey TC (1998) Immune maladaptation in the etiology of preeclampsia: a review of corroborative epidemiologic studies. Obstet Gynecol Surv 53(6):377–382PubMed
Denison FC, Grant VE, Calder AA, Kelly RW (1999) Seminal plasma components stimulate interleukin-8 and interleukin-10 release. Mol Hum Reprod 5(3):220–226PubMed
Evans CH, Lee TS, Flugelman AA (1995) Spermine-directed immunosuppression of cervical carcinoma cell sensitivity to a majority of lymphokine-activated killer lymphocyte cytotoxicity. Nat Immun 14(3):157–163PubMed
Friedline RH, Brown DS, Nguyen H, Kornfeld H, Lee J, Zhang Y, Appleby M, Der SD, Kang J, Chambers CA (2009) CD4+ regulatory T cells require CTLA-4 for the maintenance of systemic tolerance. J Exp Med 206(2):421–434. doi:10.1084/jem.20081811PubMedCentralPubMed
Guerin LR, Moldenhauer LM, Prins JR, Bromfield JJ, Hayball JD, Robertson SA (2011) Seminal fluid regulates accumulation of FOXP3+ regulatory T cells in the preimplantation mouse uterus through expanding the FOXP3+ cell pool and CCL19-mediated recruitment. Biol Reprod 85(2):397–408. doi:10.1095/biolreprod.110.088591PubMed
Gutsche S, von Wolff M, Strowitzki T, Thaler CJ (2003) Seminal plasma induces mRNA expression of IL-1beta, IL-6 and LIF in endometrial epithelial cells in vitro. Mol Hum Reprod 9(12):785–791PubMed
Hara M, Kingsley CI, Niimi M, Read S, Turvey SE, Bushell AR, Morris PJ, Powrie F, Wood KJ (2001) IL-10 is required for regulatory T cells to mediate tolerance to alloantigens in vivo. J Immunol 166(6):3789–3796PubMed
Heikkinen J, Möttönen M, Alanen A, Lassila O (2004) Phenotypic characterization of regulatory T cells in the human decidua. Clin Exp Immunol 136(2):373–378. doi:10.1111/j.1365-2249.2004.02441.xPubMedCentralPubMed
Inada K, Shima T, Nakashima A, Aoki K, Ito M, Saito S (2013) Characterization of regulatory T cells in decidua of miscarriage cases with abnormal or normal fetal chromosomal content. J Reprod Immunol 97(1):104–111. doi:10.1016/j.jri.2012.12.001PubMed
Jin L, Chen Q, Zhang T, Guo P, Li D (2009) The CD4+CD25 bright regulatory T cells and CTLA-4 expression in peripheral and decidual lymphocytes are down-regulated in human miscarriage. Clin Immunol 133(3):402–410. doi:10.1016/j.clim.2009.08.009PubMed
Juyena NS, Stelletta C (2012) Seminal plasma: an essential attribute to spermatozoa. J Androl 33(4):536–551. doi:10.2164/jandrol.110.012583PubMed
Kahn DA, Baltimore D (2010) Pregnancy induces a fetal antigen-specific maternal T regulatory cell response that contributes to tolerance. Proc Natl Acad Sci 107(20):9299–9304. doi:10.1073/pnas.1003909107PubMedCentralPubMed
Kallikourdis M, Betz AG (2007) Periodic accumulation of regulatory T cells in the uterus: preparation for the implantation of a semi-allogeneic fetus? PLoS One 2(4), e382. doi:10.1371/journal.pone.0000382PubMedCentralPubMed
Kallikourdis M, Andersen KG, Welch KA, Betz AG (2007) Alloantigen-enhanced accumulation of CCR5+ ‘effector’ regulatory T cells in the gravid uterus. Proc Natl Acad Sci USA 104(2):594–599. doi:10.1073/pnas.0604268104PubMedCentralPubMed
Kelly RW (1995) Immunosuppressive mechanisms in semen: implications for contraception. Hum Reprod 10(7):1686–1693PubMed
Kelly RW, Carr GG, Critchley HO (1997) A cytokine switch induced by human seminal plasma: an immune modulation with implications for sexually transmitted disease. Hum Reprod 12(4):677–681PubMed
Khosrotehrani K, Johnson KL, Guégan S, Stroh H, Bianchi DW (2005) Natural history of fetal cell microchimerism during and following murine pregnancy. J Reprod Immunol 66(1):1–12. doi:10.1016/j.jri.2005.02.001PubMed
Kisielewicz A, Schaier M, Schmitt E, Hug F, Haensch GM, Meuer S, Zeier M, Sohn C, Steinborn A (2010) A distinct subset of HLA-DR+-regulatory T cells is involved in the induction of preterm labor during pregnancy and in the induction of organ rejection after transplantation. Clin Immunol 137(2):209–220. doi:10.1016/j.clim.2010.07.008PubMed
Liabakk NB, Lien E, Sundan A, Sunde A, Austgulen R, Espevik T (1993) High concentrations of the soluble p55 tumour necrosis factor receptor in human seminal plasma. Hum Reprod 8(11):1837–1842PubMed
Lim HW, Hillsamer P, Banham AH, Kim CH (2005) Cutting edge: direct suppression of B cells by CD4+ CD25+ regulatory T cells. J Immunol 175(7):4180–4183PubMed
Liu C, Wang X, Sun X (2013) Assessment of sperm antigen specific T regulatory cells in women with recurrent miscarriage. Early Hum Dev 89(2):95–100. doi:10.1016/j.earlhumdev.2012.08.003PubMed
Lokeshwar BL, Block NL (1992) Isolation of a prostate carcinoma cell proliferation-inhibiting factor from human seminal plasma and its similarity to transforming growth factor beta. Cancer Res 52(20):5821–5825PubMed
Loubière LS, Lambert NC, Flinn LJ, Erickson TD, Yan Z, Guthrie KA, Vickers KT, Nelson JL (2006) Maternal microchimerism in healthy adults in lymphocytes, monocyte/macrophages and NK cells. Lab Invest 86(11):1185–1192. doi:10.1038/labinvest.3700471PubMed
McMaster MT, Newton RC, Dey SK, Andrews GK (1992) Activation and distribution of inflammatory cells in the mouse uterus during the preimplantation period. J Immunol 148(6):1699–1705PubMed
Mei S, Tan J, Chen H, Chen Y, Zhang J (2010) Changes of CD4+CD25high regulatory T cells and FOXP3 expression in unexplained recurrent spontaneous abortion patients. Fertil Steril 94(6):2244–2247. doi:10.1016/j.fertnstert.2009.11.020PubMed
Mempel TR, Pittet MJ, Khazaie K, Weninger W, Weissleder R, von Boehmer H, von Andrian UH (2006) Regulatory T cells reversibly suppress cytotoxic T cell function independent of effector differentiation. Immunity 25(1):129–141. doi:10.1016/j.immuni.2006.04.015PubMed
Milardi D, Grande G, Vincenzoni F, Castagnola M, Marana R (2013) Proteomics of human seminal plasma: identification of biomarker candidates for fertility and infertility and the evolution of technology. Mol Reprod Dev 80(5):350–357. doi:10.1002/mrd.22178PubMed
Misra N, Bayry J, Lacroix-Desmazes S, Kazatchkine MD, Kaveri SV (2004) Cutting edge: human CD4+CD25+ T cells restrain the maturation and antigen-presenting function of dendritic cells. J Immunol 172(8):4676–4680PubMed
Mjösberg J, Berg G, Ernerudh J, Ekerfelt C (2007) CD4+ CD25+ regulatory T cells in human pregnancy: development of a Treg-MLC-ELISPOT suppression assay and indications of paternal specific Tregs. Immunology 120(4):456–466. doi:10.1111/j.1365-2567.2006.02529.xPubMedCentralPubMed
Moldenhauer LM, Diener KR, Thring DM, Brown MP, Hayball JD, Robertson SA (2009) Cross-presentation of male seminal fluid antigens elicits T cell activation to initiate the female immune response to pregnancy. J Immunol 182(12):8080–8093. doi:10.4049/jimmunol.0804018PubMed
Mori A, Zhai YL, Toki T, Nikaido T, Fujii S (1997) Distribution and heterogeneity of mast cells in the human uterus. Hum Reprod 12(2):368–372PubMed
Nocera M, Chu TM (1993) Transforming growth factor beta as an immunosuppressive protein in human seminal plasma. Am J Reprod Immunol 30(1):1–8PubMed
Nocera M, Chu TM (1995) Characterization of latent transforming growth factor-beta from human seminal plasma. Am J Reprod Immunol 33(4):282–291PubMed
O’Leary S, Armstrong DT, Robertson SA (2013) Transforming growth factor-? (TGF?) in porcine seminal plasma. Reprod Fertil Dev 23(6):748. doi:10.1071/RD11001
Padilla L, Reinicke K, Montesino H, Villena F, Asencio H, Cruz M, Rudolph MI (1990) Histamine content and mast cells distribution in mouse uterus: the effect of sexual hormones, gestation and labor. Cell Mol Biol 36(1):93–100PubMed
Pakravan N, Ghaffarinia A, Jalili C, Riazi-Rad F, Tajedini M, Mostafaie A (2014) Seminal vesicle fluid ameliorates experimental autoimmune encephalomyelitis by increasing FoxP3+ regulatory T-cell numbers. Cell Mol Immunol. doi:10.1038/cmi.2014.88PubMed
Pandya IJ, Cohen J (1985) The leukocytic reaction of the human uterine cervix to spermatozoa. Fertil Steril 43(3):417–421PubMed
Piccirillo CA, Shevach EM (2001) Cutting edge: control of CD8+ T cell activation by CD4+CD25+ immunoregulatory cells. J Immunol 167(3):1137–1140PubMed
Plaks V, Birnberg T, Berkutzki T, Sela S, BenYashar A, Kalchenko V, Mor G, Keshet E, Dekel N, Neeman M, Jung S (2008) Uterine DCs are crucial for decidua formation during embryo implantation in mice. J Clin Invest. doi:10.1172/JCI36682PubMedCentralPubMed
Prins JR, Boelens HM, Heimweg J, van der Heide S, Dubois AE, van Oosterhout AJ, Erwich JJHM (2009) Preeclampsia is associated with lower percentages of regulatory T cells in maternal blood. Hypertens Pregnancy 28(3):300–311. doi:10.1080/10641950802601237PubMed
Quinn KH, Lacoursiere DY, Cui L, Bui J, Parast MM (2011) The unique pathophysiology of early-onset severe preeclampsia: role of decidual T regulatory cells. J Reprod Immunol 91(1–2):76–82. doi:10.1016/j.jri.2011.05.006PubMed
Ramhorst R, Fraccaroli L, Aldo P, Alvero AB, Cardenas I, Leirós CP, Mor G (2012) Modulation and recruitment of inducible regulatory T cells by first trimester trophoblast cells. Am J Reprod Immunol 67(1):17–27. doi:10.1111/j.1600-0897.2011.01056.xPubMedCentralPubMed
Robertson SA (2005) Seminal plasma and male factor signalling in the female reproductive tract. Cell Tissue Res 322(1):43–52. doi:10.1007/s00441-005-1127-3PubMed
Robertson SA, Mau VJ, Tremellen KP, Seamark RF (1996) Role of high molecular weight seminal vesicle proteins in eliciting the uterine inflammatory response to semen in mice. J Reprod Fertil 107(2):265–277PubMed
Robertson SA, Guerin LR, Bromfield JJ, Branson KM, Ahlström AC, Care AS (2009) Seminal fluid drives expansion of the CD4+CD25+ T regulatory cell pool and induces tolerance to paternal alloantigens in mice. Biol Reprod 80(5):1036–1045. doi:10.1095/biolreprod.108.074658PubMedCentralPubMed
Root-Bernstein RS, DeWitt SH (1995) Semen alloantigens and lymphocytotoxic antibodies in AIDS and ICL. Genetica 95(1–3):133–156PubMed
Rowe JH, Ertelt JM, Xin L, Way SS (2012) Pregnancy imprints regulatory memory that sustains anergy to fetal antigen. Nature 490(7418):102–106. doi:10.1038/nature11462PubMedCentralPubMed
Saftlas AF, Rubenstein L, Prater K, Harland KK, Field E, Triche EW (2014) Cumulative exposure to paternal seminal fluid prior to conception and subsequent risk of preeclampsia. J Reprod Immunol 101–102:104–110. doi:10.1016/j.jri.2013.07.006PubMed
Saito S, Sasaki Y, Sakai M (2005) CD4(+)CD25high regulatory T cells in human pregnancy. J Reprod Immunol 65(2):111–120. doi:10.1016/j.jri.2005.01.004PubMed
Sakaguchi S, Sakaguchi N, Asano M, Itoh M, Toda M (1995) Immunologic self-tolerance maintained by activated T cells expressing IL-2 receptor alpha-chains (CD25). Breakdown of a single mechanism of self-tolerance causes various autoimmune diseases. J Immunol 155(3):1151–1164PubMed
Santner-Nanan B, Peek MJ, Khanam R, Richarts L, Zhu E, de St F, Groth B, Nanan R (2009) Systemic increase in the ratio between Foxp3+ and IL-17-producing CD4+ T cells in healthy pregnancy but not in preeclampsia. J Immunol 183(11):7023–7030. doi:10.4049/jimmunol.0901154PubMed
Sasaki Y, Sakai M, Miyazaki S, Higuma S, Shiozaki A, Saito S (2004) Decidual and peripheral blood CD4+CD25+ regulatory T cells in early pregnancy subjects and spontaneous abortion cases. Mol Hum Reprod 10(5):347–353. doi:10.1093/molehr/gah044PubMed
Sasaki Y, Darmochwal-Kolarz D, Suzuki D, Sakai M, Ito M, Shima T, Shiozaki A, Rolinski J, Saito S (2007) Proportion of peripheral blood and decidual CD4(+) CD25(bright) regulatory T cells in pre-eclampsia. Clin Exp Immunol 149(1):139–145. doi:10.1111/j.1365-2249.2007.03397.xPubMedCentralPubMed
Schober L, Radnai D, Schmitt E, Mahnke K, Sohn C, Steinborn A (2012) Term and preterm labor: decreased suppressive activity and changes in composition of the regulatory T-cell pool. Immunol Cell Biol 90(10):935–944. doi:10.1038/icb.2012.33PubMed
Schumacher A, Wafula PO, Bertoja AZ, Sollwedel A, Thuere C, Wollenberg I, Yagita H, Volk H, Zenclussen AC (2007) Mechanisms of action of regulatory T cells specific for paternal antigens during pregnancy. Obstet Gynecol 110(5):1137–1145. doi:10.1097/01.AOG.0000284625.10175.31PubMed
Schumacher A, Brachwitz N, Sohr S, Engeland K, Langwisch S, Dolaptchieva M, Alexander T, Taran A, Malfertheiner SF, Costa S, Zimmermann G, Nitschke C, Volk H, Alexander H, Gunzer M, Zenclussen AC (2009) Human chorionic gonadotropin attracts regulatory T cells into the fetal-maternal interface during early human pregnancy. J Immunol 182(9):5488–5497. doi:10.4049/jimmunol.0803177PubMed
Schumacher A, Wafula PO, Teles A, El-Mousleh T, Linzke N, Zenclussen ML, Langwisch S, Heinze K, Wollenberg I, Casalis PA, Volk H, Fest S, Zenclussen AC, Kassiotis G (2012) Blockage of heme oxygenase-1 abrogates the protective effect of regulatory T cells on murine pregnancy and promotes the maturation of dendritic cells. PLoS One 7(8), e42301. doi:10.1371/journal.pone.0042301PubMedCentralPubMed
Schumacher A, Heinze K, Witte J, Poloski E, Linzke N, Woidacki K, Zenclussen AC (2013) Human chorionic gonadotropin as a central regulator of pregnancy immune tolerance. J Immunol 190(6):2650–2658. doi:10.4049/jimmunol.1202698PubMed
Schumacher A, Poloski E, Spörke D, Zenclussen AC (2014) Luteinizing hormone contributes to fetal tolerance by regulating adaptive immune responses. Am J Reprod Immunol 71(5):434–440. doi:10.1111/aji.12215PubMed
Seol H, Oh M, Lim J, Jung N, Yoon S, Kim H (2008) The role of CD4+CD25bright regulatory T cells in the maintenance of pregnancy, premature rupture of membranes, and labor. Yonsei Med J 49(3):366. doi:10.3349/ymj.2008.49.3.366PubMedCentralPubMed
Sharkey DJ, Macpherson AM, Tremellen KP, Robertson SA (2007) Seminal plasma differentially regulates inflammatory cytokine gene expression in human cervical and vaginal epithelial cells. Mol Hum Reprod 13(7):491–501. doi:10.1093/molehr/gam028PubMed
Slater M, Murphy CR (1999) Thrombospondin is sequentially expressed and then de-expressed during early pregnancy in the rat uterus. Histochem J 31(7):471–475PubMed
Somerset DA, Zheng Y, Kilby MD, Sansom DM, Drayson MT (2004) Normal human pregnancy is associated with an elevation in the immune suppressive CD25+ CD4+ regulatory T-cell subset. Immunology 112(1):38–43. doi:10.1111/j.1365-2567.2004.01869.xPubMedCentralPubMed
Srivastava MD, Lippes J, Srivastava BI (1996) Cytokines of the human reproductive tract. Am J Reprod Immunol 36(3):157–166PubMed
Stewart CL, Kaspar P, Brunet LJ, Bhatt H, Gadi I, Köntgen F, Abbondanzo SJ (1992) Blastocyst implantation depends on maternal expression of leukaemia inhibitory factor. Nature 359(6390):76–79. doi:10.1038/359076a0PubMed
Taams LS, van Amelsfort JMR, Tiemessen MM, Jacobs KMG, de Jong EC, Akbar AN, Bijlsma JWJ, Lafeber FPJG (2005) Modulation of monocyte/macrophage function by human CD4+CD25+ regulatory T cells. Hum Immunol 66(3):222–230. doi:10.1016/j.humimm.2004.12.006PubMedCentralPubMed
Tan X, Liao H, Sun L, Okabe M, Xiao Z, Dawe GS (2005) Fetal microchimerism in the maternal mouse brain: a novel population of fetal progenitor or stem cells able to cross the blood-brain barrier? Stem Cells 23(10):1443–1452. doi:10.1634/stemcells.2004-0169PubMed
Tarter TH, Alexander NJ (1984) Complement-inhibiting activity of seminal plasma. Am J Reprod Immunol 6(1):28–32PubMed
Teles A, Schumacher A, Kühnle M, Linzke N, Thuere C, Reichardt P, Tadokoro CE, Hämmerling GJ, Zenclussen AC (2013a) Control of uterine microenvironment by Foxp3+ cells facilitates embryo implantation. Front Immunol 4:158. doi:10.3389/fimmu.2013.00158PubMedCentralPubMed
Teles A, Thuere C, Wafula PO, El-Mousleh T, Zenclussen ML, Zenclussen AC (2013b) Origin of Foxp3(+) cells during pregnancy. Am J Clin Exp Immunol 2(3):222–233PubMedCentralPubMed
Thuere C, Zenclussen ML, Schumacher A, Langwisch S, Schulte-Wrede U, Teles A, Paeschke S, Volk H, Zenclussen AC (2007) Kinetics of regulatory T cells during murine pregnancy. Am J Reprod Immunol 58(6):514–523. doi:10.1111/j.1600-0897.2007.00538.xPubMed
Tilburgs T, Roelen DL, van der Mast BJ, van Schip JJ, Kleijburg C, de Groot-Swings GM, Kanhai HHH, Claas FHJ, Scherjon SA (2006) Differential distribution of CD4(+)CD25(bright) and CD8(+)CD28(-) T-cells in decidua and maternal blood during human pregnancy. Placenta 27:S47–S53. doi:10.1016/j.placenta.2005.11.008PubMed
Tilburgs T, Roelen DL, van der Mast BJ, de Groot-Swings GM, Kleijburg C, Scherjon SA, Claas FH (2008) Evidence for a selective migration of fetus-specific CD4+CD25bright regulatory T cells from the peripheral blood to the decidua in human pregnancy. J Immunol 180(8):5737–5745PubMed
Tilburgs T, Scherjon SA, van der Mast BJ, Haasnoot GW, Versteeg-V D, Voort-Maarschalk M, Roelen DL, van Rood JJ, Claas FHJ (2009) Fetal-maternal HLA-C mismatch is associated with decidual T cell activation and induction of functional T regulatory cells. J Reprod Immunol 82(2):148–157. doi:10.1016/j.jri.2009.05.003PubMed
Toldi G, Švec P, Vásárhelyi B, Mészáros G, Rigó J, Tulassay T, Treszl A (2008) Decreased number of FoxP3+ regulatory T cells in preeclampsia. Acta Obstet Gynecol Scand 87(11):1229–1233. doi:10.1080/00016340802389470PubMed
Tremellen KP, Seamark RF, Robertson SA (1998) Seminal transforming growth factor beta1 stimulates granulocyte-macrophage colony-stimulating factor production and inflammatory cell recruitment in the murine uterus. Biol Reprod 58(5):1217–1225PubMed
Verdijk RM, Kloosterman A, Pool J, van de Keur M, Naipal AMIH, van Halteren AGS, Brand A, Mutis T, Goulmy E (2004) Pregnancy induces minor histocompatibility antigen-specific cytotoxic T cells: implications for stem cell transplantation and immunotherapy. Blood 103(5):1961–1964. doi:10.1182/blood-2003-05-1625PubMed
Wahl SM, Swisher J, McCartney-Francis N, Chen W (2004) TGF-beta: the perpetrator of immune suppression by regulatory T cells and suicidal T cells. J Leukoc Biol 76(1):15–24. doi:10.1189/jlb.1103539PubMed
Weinberg A, Enomoto L, Marcus R, Canniff J (2011) Effect of menstrual cycle variation in female sex hormones on cellular immunity and regulation. J Reprod Immunol 89(1):70–77. doi:10.1016/j.jri.2010.11.009PubMed
Witkin SS, Goldstein M (1988) Reduced levels of T suppressor/cytotoxic lymphocytes in semen from vasovasostomized men: relationship to sperm autoantibodies. J Reprod Immunol 14(3):283–290PubMed
Woidacki K, Popovic M, Metz M, Schumacher A, Linzke N, Teles A, Poirier F, Fest S, Jensen F, Rabinovich GA, Maurer M, Zenclussen AC (2013) Mast cells rescue implantation defects caused by c-kit deficiency. Cell Death Dis 4(1):e462. doi:10.1038/cddis.2012.214PubMedCentralPubMed
Wu L, Luo LH, Zhang YX, Li Q, Xu B, Zhou GX, Luan HB, Liu YS (2014) Alteration of Th17 and Treg cells in patients with unexplained recurrent spontaneous abortion before and after lymphocyte immunization therapy. Reprod Biol Endocrinol 12:74. doi:10.1186/1477-7827-12-74PubMedCentralPubMed
Xiong H, Zhou C, Qi G (2010) Proportional changes of CD4+CD25+Foxp3+ regulatory T cells in maternal peripheral blood during pregnancy and labor at term and preterm. Clin Invest Med 33(6):E422PubMed
Xiong Y, Yuan Z, He L (2013) Effects of estrogen on CD4(+) CD25(+) regulatory T cell in peripheral blood during pregnancy. Asian Pac J Trop Med 6(9):748–752. doi:10.1016/S1995-7645(13)60131-5PubMed
Yan Z, Lambert NC, Guthrie KA, Porter AJ, Loubiere LS, Madeleine MM, Stevens AM, Hermes HM, Nelson JL (2005) Male microchimerism in women without sons: quantitative assessment and correlation with pregnancy history. Am J Med 118(8):899–906. doi:10.1016/j.amjmed.2005.03.037PubMed
Yang H, Qiu L, Chen G, Ye Z, Lü C, Lin Q (2008) Proportional change of CD4+CD25+ regulatory T cells in decidua and peripheral blood in unexplained recurrent spontaneous abortion patients. Fertil Steril 89(3):656–661. doi:10.1016/j.fertnstert.2007.03.037PubMed
Yin Y, Han X, Shi Q, Zhao Y, He Y (2012) Adoptive transfer of CD4+CD25+ regulatory T cells for prevention and treatment of spontaneous abortion. Eur J Obstet Gynecol Reprod Biol 161(2):177–181. doi:10.1016/j.ejogrb.2011.12.023PubMed
Zenclussen AC, Gerlof K, Zenclussen ML, Sollwedel A, Bertoja AZ, Ritter T, Kotsch K, Leber J, Volk H (2005) Abnormal T-cell reactivity against paternal antigens in spontaneous abortion: adoptive transfer of pregnancy-induced CD4+CD25+ T regulatory cells prevents fetal rejection in a murine abortion model. Am J Pathol 166(3):811–822. doi:10.1016/S0002-9440(10)62302-4PubMedCentralPubMed
Zenclussen ML, Thuere C, Ahmad N, Wafula PO, Fest S, Teles A, Leber A, Casalis PA, Bechmann I, Priller J, Volk H, Zenclussen AC (2010) The persistence of paternal antigens in the maternal body is involved in regulatory T-cell expansion and fetal-maternal tolerance in murine pregnancy. Am J Reprod Immunol 63(3):200–208. doi:10.1111/j.1600-0897.2009.00793.xPubMed
Zenclussen AC, Olivieri DN, Dustin ML, Tadokoro CE (2013) In vivo multiphoton microscopy technique to reveal the physiology of the mouse uterus. Am J Reprod Immunol 69(3):281–289. doi:10.1111/aji.12066PubMed
Zhou J, Wang Z, Zhao X, Wang J, Sun H, Hu Y (2012) An increase of Treg cells in the peripheral blood is associated with a better in vitro fertilization treatment outcome. Am J Reprod Immunol 68(2):100–106. doi:10.1111/j.1600-0897.2012.01153.xPubMed